

Beyond the Exome: The Non-coding Genome and Enhancers in Neurodevelopmental Disorders and Malformations of Cortical Development
- PMID: 31417368
- PMCID: PMC6685065
- DOI: 10.3389/fncel.2019.00352
Beyond the Exome: The Non-coding Genome and Enhancers in Neurodevelopmental Disorders and Malformations of Cortical Development
Abstract
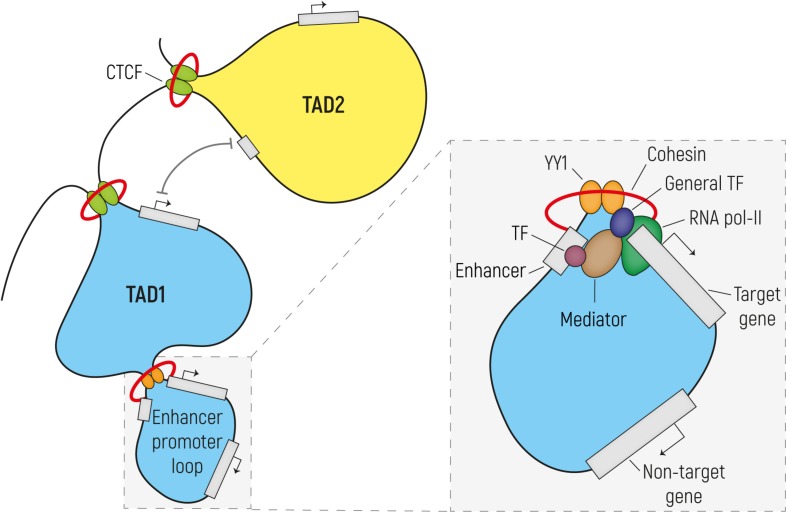
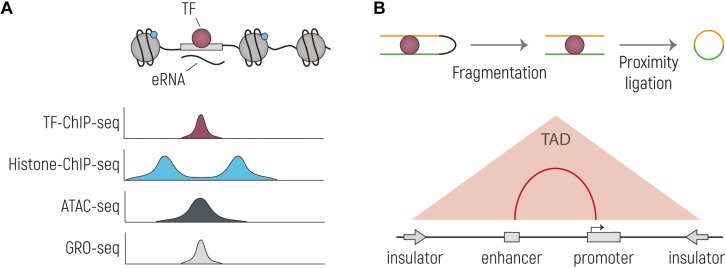
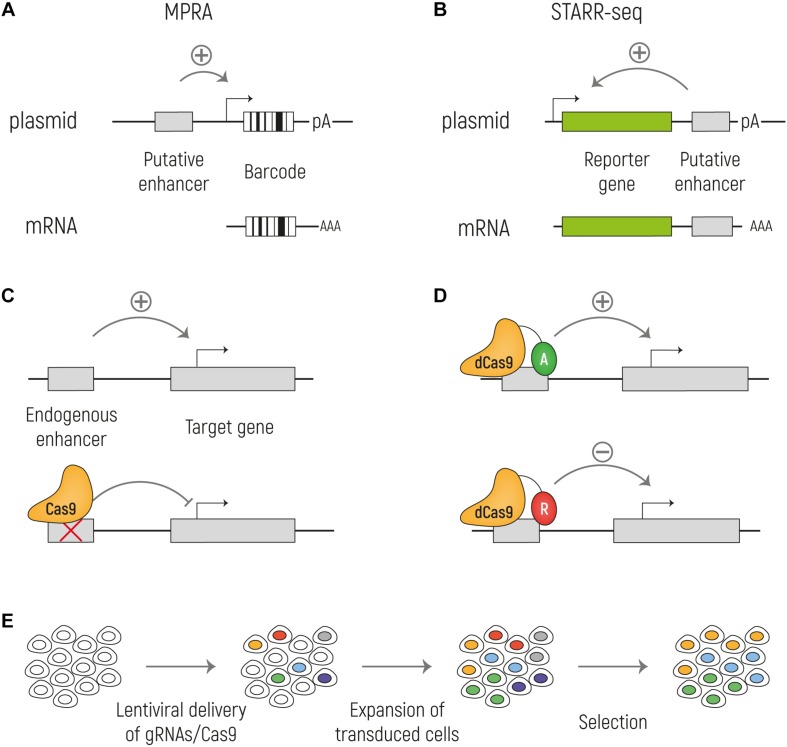
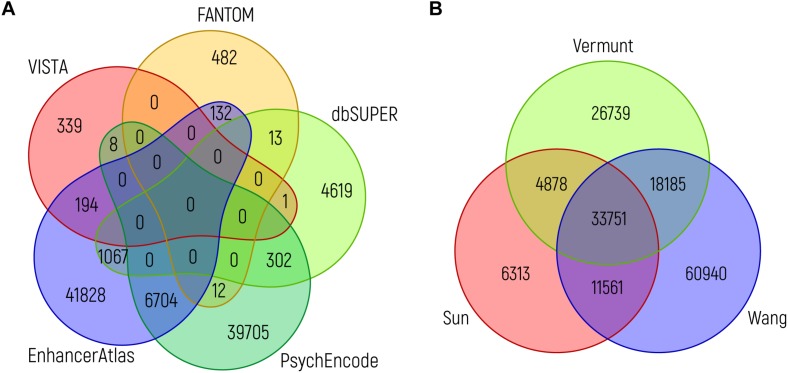
The development of the human cerebral cortex is a complex and dynamic process, in which neural stem cell proliferation, neuronal migration, and post-migratory neuronal organization need to occur in a well-organized fashion. Alterations at any of these crucial stages can result in malformations of cortical development (MCDs), a group of genetically heterogeneous neurodevelopmental disorders that present with developmental delay, intellectual disability and epilepsy. Recent progress in genetic technologies, such as next generation sequencing, most often focusing on all protein-coding exons (e.g., whole exome sequencing), allowed the discovery of more than a 100 genes associated with various types of MCDs. Although this has considerably increased the diagnostic yield, most MCD cases remain unexplained. As Whole Exome Sequencing investigates only a minor part of the human genome (1-2%), it is likely that patients, in which no disease-causing mutation has been identified, could harbor mutations in genomic regions beyond the exome. Even though functional annotation of non-coding regions is still lagging behind that of protein-coding genes, tremendous progress has been made in the field of gene regulation. One group of non-coding regulatory regions are enhancers, which can be distantly located upstream or downstream of genes and which can mediate temporal and tissue-specific transcriptional control via long-distance interactions with promoter regions. Although some examples exist in literature that link alterations of enhancers to genetic disorders, a widespread appreciation of the putative roles of these sequences in MCDs is still lacking. Here, we summarize the current state of knowledge on cis-regulatory regions and discuss novel technologies such as massively-parallel reporter assay systems, CRISPR-Cas9-based screens and computational approaches that help to further elucidate the emerging role of the non-coding genome in disease. Moreover, we discuss existing literature on mutations or copy number alterations of regulatory regions involved in brain development. We foresee that the future implementation of the knowledge obtained through ongoing gene regulation studies will benefit patients and will provide an explanation to part of the missing heritability of MCDs and other genetic disorders.
Keywords: bioinformatics; cis-regulatory elements; enhancers; epigenome; functional genomics; gene regulation; malformations of cortical development; massively parallel reporter assays.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials