

CD8αα homodimers function as a coreceptor for KIR3DL1
- PMID: 31420518
- PMCID: PMC6731649
- DOI: 10.1073/pnas.1905943116
CD8αα homodimers function as a coreceptor for KIR3DL1
Abstract
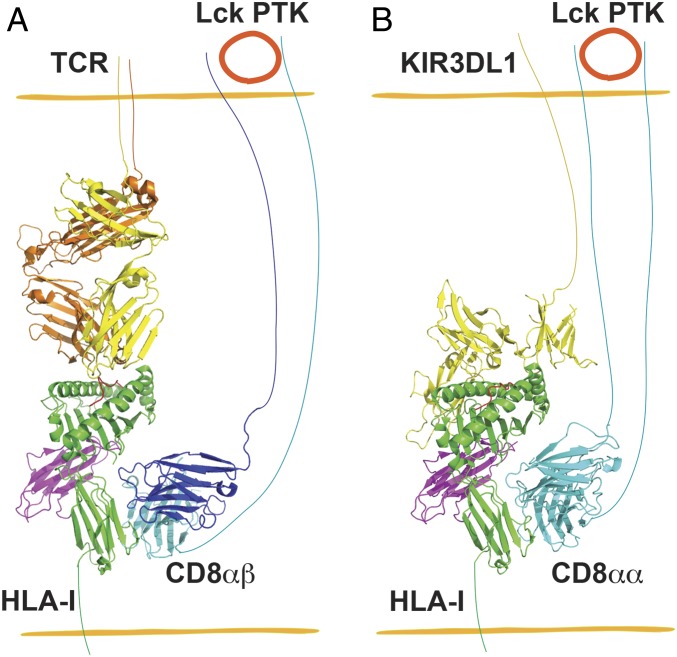
Cluster of differentiation 8 (CD8) is a cell surface glycoprotein, which is expressed as 2 forms, αα homodimer or αβ heterodimer. Peptide-loaded major histocompatibility complex class I (pMHC-I) molecules are major ligands for both forms of CD8. CD8αβ is a coreceptor for the T cell receptor (TCR) and binds to the same cognate pMHC-I as the TCR, thus enabling or augmenting T cell responses. The function of CD8αα homodimers is largely unknown. While CD8αβ heterodimer is expressed exclusively on CD8+ T cells, the CD8αα homodimer is present in subsets of T cells and human natural killer (NK) cells. Here, we report that the CD8αα homodimer functions as a coreceptor for KIR3DL1, an inhibitory receptor of NK cells that is specific for certain MHC-I allotypes. CD8αα enhances binding of pMHC-I to KIR3DL1, increases KIR3DL1 clustering at the immunological synapse, and augments KIR3DL1-mediated inhibition of NK cell activation. Additionally, interactions between pMHC-I and CD8αα homodimers regulate KIR3DL1+ NK cell education. Together, these findings reveal another dimension to the modulation of NK cell activity.
Keywords: CD8αα; KIR3DL1; NK cell activation; coreceptor.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Cole D. K., Gao G. F., CD8: Adhesion molecule, co-receptor and immuno-modulator. Cell. Mol. Immunol. 1, 81–88 (2004). - PubMed
-
- Purbhoo M. A., et al. , The human CD8 coreceptor effects cytotoxic T cell activation and antigen sensitivity primarily by mediating complete phosphorylation of the T cell receptor ζ chain. J. Biol. Chem. 276, 32786–32792 (2001). - PubMed
-
- Garcia K. C., et al. , CD8 enhances formation of stable T-cell receptor/MHC class I molecule complexes. Nature 384, 577–581 (1996). - PubMed
-
- Huang J., Edwards L. J., Evavold B. D., Zhu C., Kinetics of MHC-CD8 interaction at the T cell membrane. J. Immunol. 179, 7653–7662 (2007). - PubMed
-
- Hayday A., Theodoridis E., Ramsburg E., Shires J., Intraepithelial lymphocytes: Exploring the third way in immunology. Nat. Immunol. 2, 997–1003 (2001). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
