

Functional Characterization of an Interferon Gamma Receptor-Like Protein on Entamoeba histolytica
- PMID: 31427448
- PMCID: PMC6803330
- DOI: 10.1128/IAI.00540-19
Functional Characterization of an Interferon Gamma Receptor-Like Protein on Entamoeba histolytica
Abstract
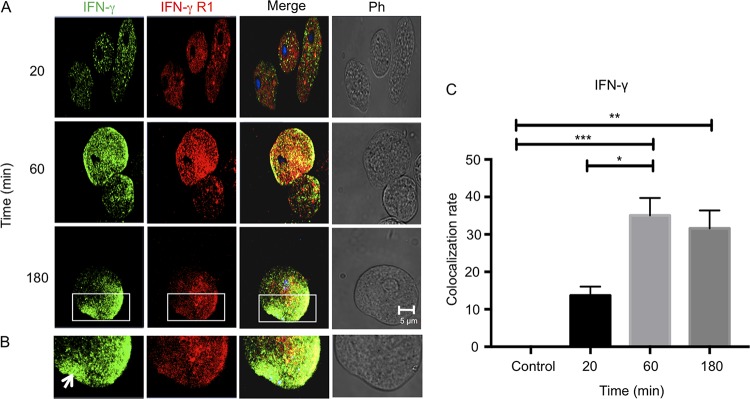
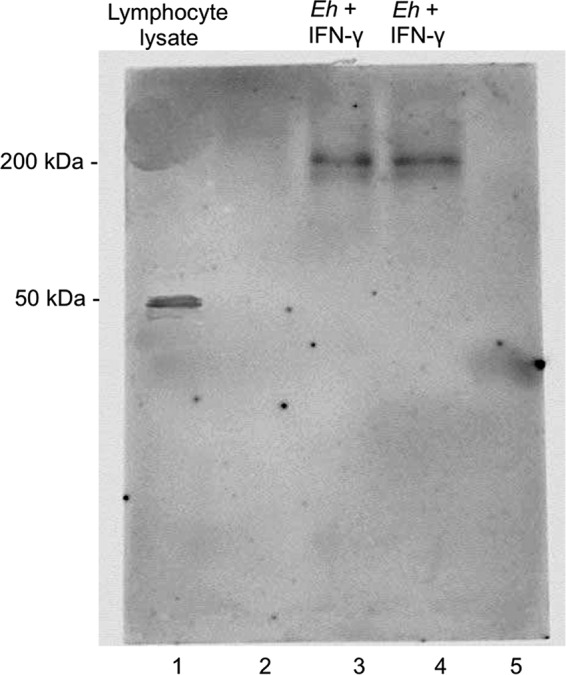
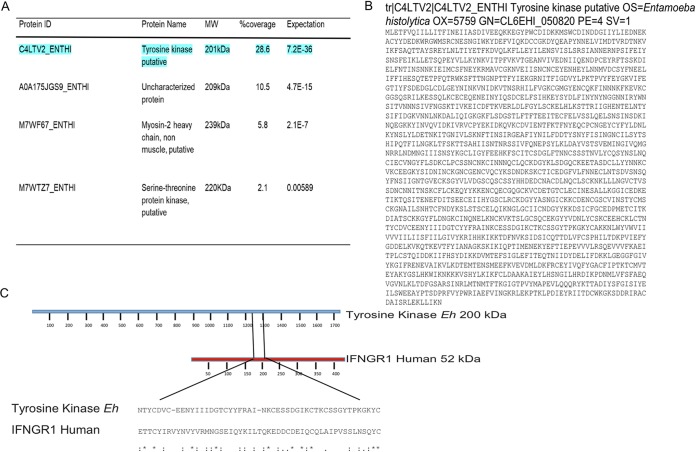
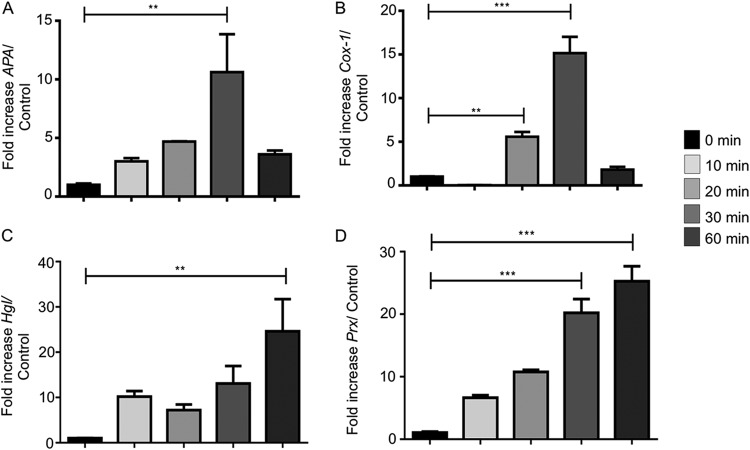
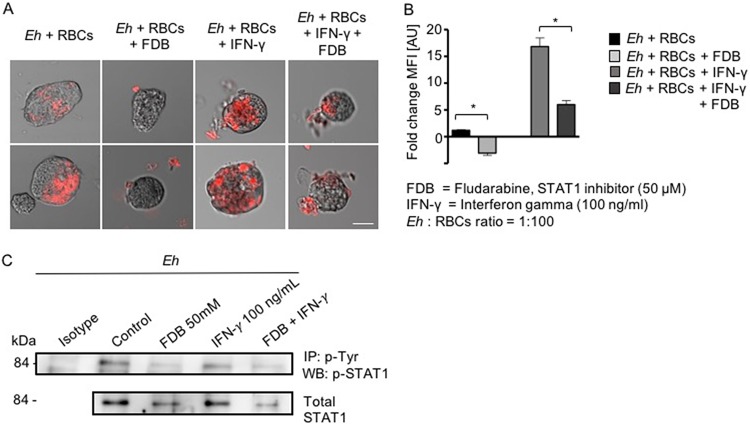
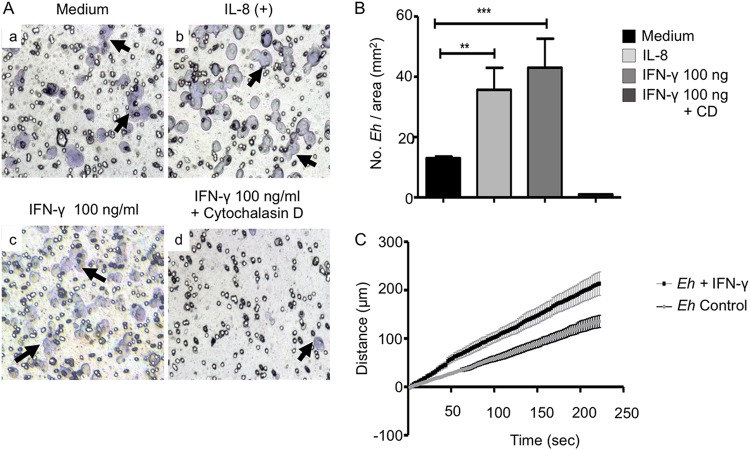
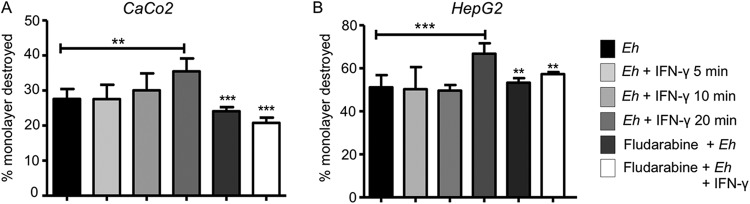
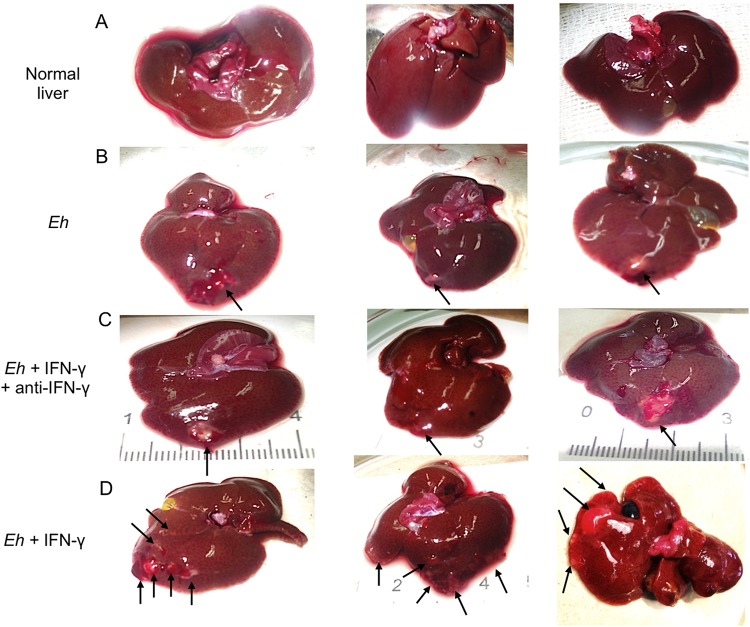
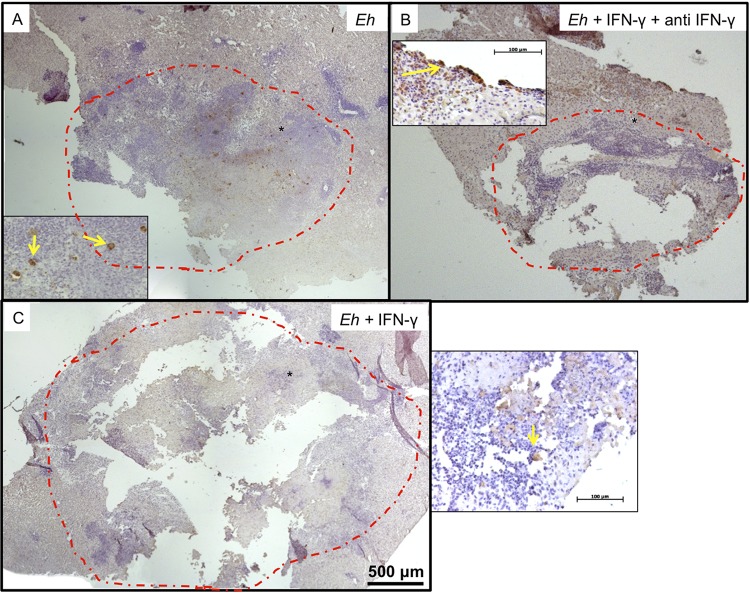
Entamoeba histolytica is an anaerobic parasitic protozoan and the causative agent of amoebiasis. E. histolytica expresses proteins that are structurally homologous to human proteins and uses them as virulence factors. We have previously shown that E. histolytica binds exogenous interferon gamma (IFN-γ) on its surface, and in this study, we explored whether exogenous IFN-γ could modulate parasite virulence. We identified an IFN-γ receptor-like protein on the surface of E. histolytica trophozoites by using anti-IFN-γ receptor 1 (IFN-γR1) antibody and performing immunofluorescence, Western blot, protein sequencing, and in silico analyses. Coupling of human IFN-γ to the IFN-γ receptor-like protein on live E. histolytica trophozoites significantly upregulated the expression of E. histolytica cysteine protease A1 (EhCP-A1), EhCP-A2, EhCP-A4, EhCP-A5, amebapore A (APA), cyclooxygenase 1 (Cox-1), Gal-lectin (Hgl), and peroxiredoxin (Prx) in a time-dependent fashion. IFN-γ signaling via the IFN-γ receptor-like protein enhanced E. histolytica's erythrophagocytosis of human red blood cells, which was abrogated by the STAT1 inhibitor fludarabine. Exogenous IFN-γ enhanced chemotaxis of E. histolytica, its killing of Caco-2 colonic and Hep G2 liver cells, and amebic liver abscess formation in hamsters. These results demonstrate that E. histolytica expresses a surface IFN-γ receptor-like protein that is functional and may play a role in disease pathogenesis and/or immune evasion.
Keywords: Entamoeba histolytica; IFN-γ; amoebiasis; cysteine proteases; cytopathic effect; erythrophagocytosis.
Copyright © 2019 Pulido-Ortega et al.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
