

Transcriptionally active HERV-H retrotransposons demarcate topologically associating domains in human pluripotent stem cells
- PMID: 31427791
- PMCID: PMC6722002
- DOI: 10.1038/s41588-019-0479-7
Transcriptionally active HERV-H retrotransposons demarcate topologically associating domains in human pluripotent stem cells
Abstract
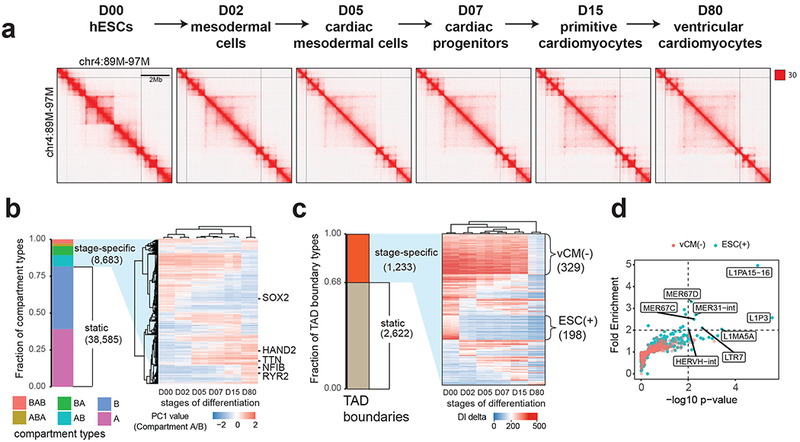
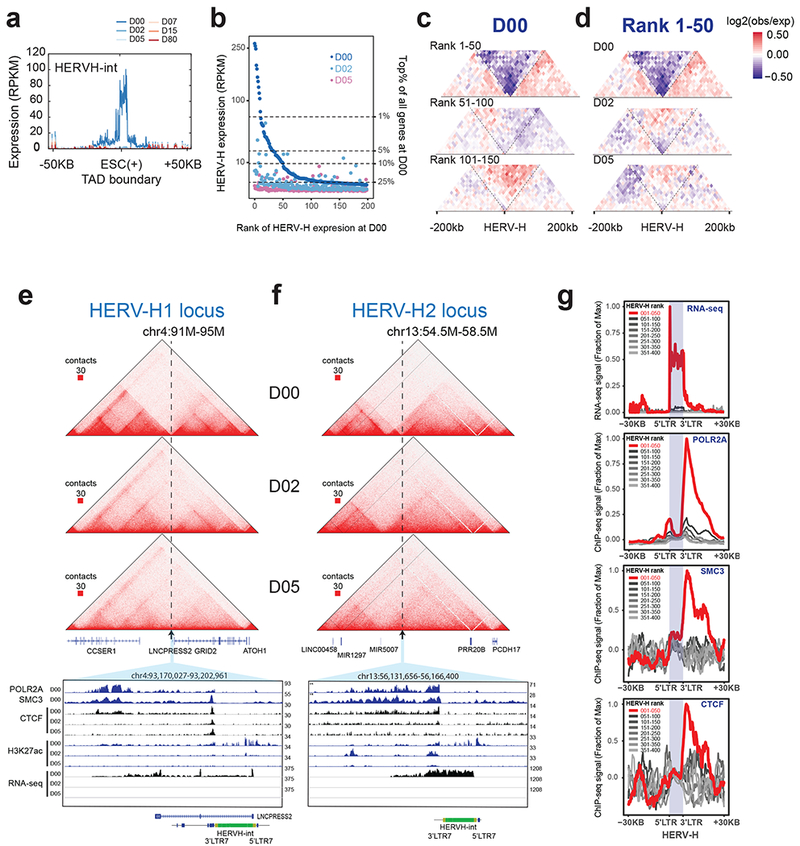
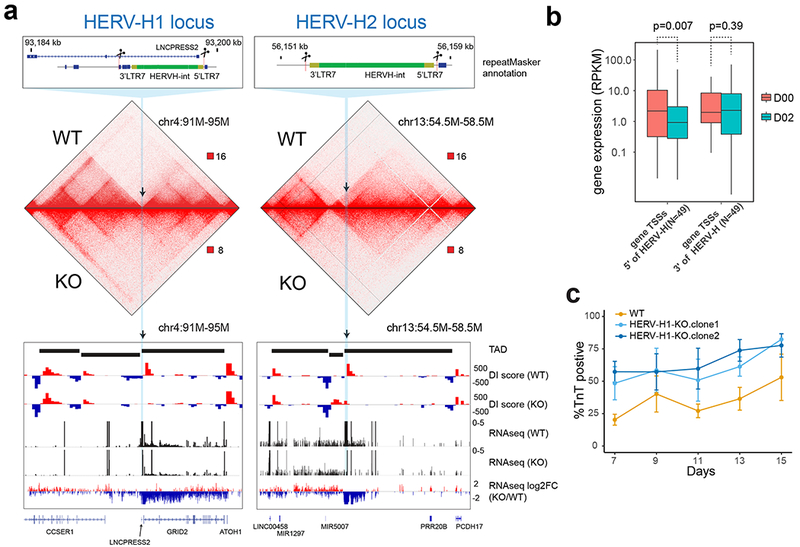
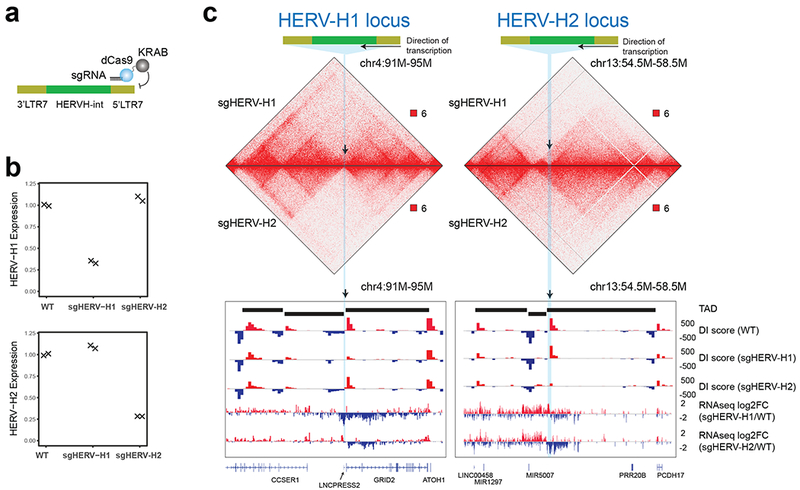
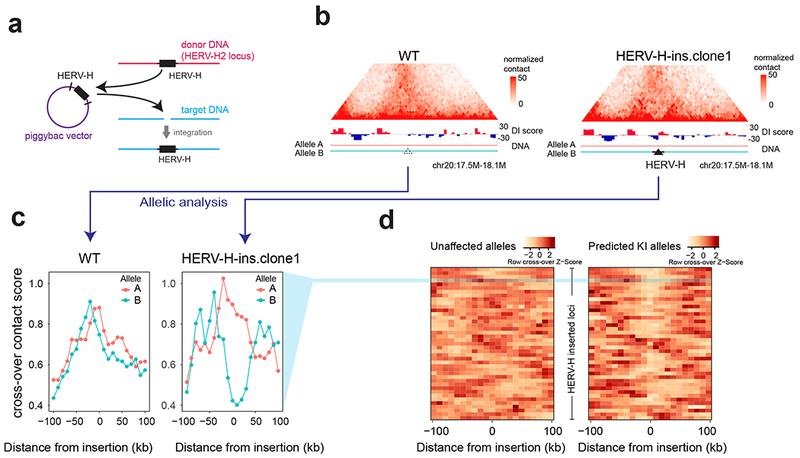
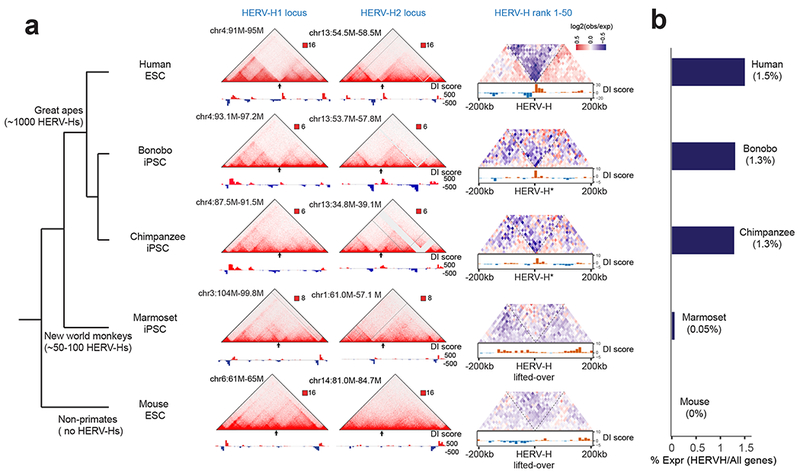
Chromatin architecture has been implicated in cell type-specific gene regulatory programs, yet how chromatin remodels during development remains to be fully elucidated. Here, by interrogating chromatin reorganization during human pluripotent stem cell (hPSC) differentiation, we discover a role for the primate-specific endogenous retrotransposon human endogenous retrovirus subfamily H (HERV-H) in creating topologically associating domains (TADs) in hPSCs. Deleting these HERV-H elements eliminates their corresponding TAD boundaries and reduces the transcription of upstream genes, while de novo insertion of HERV-H elements can introduce new TAD boundaries. The ability of HERV-H to create TAD boundaries depends on high transcription, as transcriptional repression of HERV-H elements prevents the formation of boundaries. This ability is not limited to hPSCs, as these actively transcribed HERV-H elements and their corresponding TAD boundaries also appear in pluripotent stem cells from other hominids but not in more distantly related species lacking HERV-H elements. Overall, our results provide direct evidence for retrotransposons in actively shaping cell type- and species-specific chromatin architecture.
Conflict of interest statement
Competing Interests Statement:
B.R. is a co-founder of Arima Genomics, Inc.
Figures
References
-
- Dekker J & Mirny L The 3D Genome as Moderator of Chromosomal Communication. Cell 164, 1110–1121, doi:10.1016/j.cell.2016.02.007 (2016). - DOI - PMC - PubMed
-
- Yu M & Ren B The Three-Dimensional Organization of Mammalian Genomes. Annu Rev Cell Dev Biol 33, 265–289, doi:10.1146/annurev-cellbio-100616-060531 (2017). - DOI - PMC - PubMed
-
- Meaburn KJ & Misteli T Cell biology: chromosome territories. Nature 445, 379–781, doi:10.1038/445379a (2007). - DOI - PubMed
-
- Lieberman-Aiden E et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science 326, 289–293, doi:10.1126/science.1181369 (2009). - DOI - PMC - PubMed
-
- Dixon JR et al. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature 485, 376–380, doi:10.1038/nature11082 (2012). - DOI - PMC - PubMed
Methods-only references
-
- Lian X et al. Directed cardiomyocyte differentiation from human pluripotent stem cells by modulating Wnt/beta-catenin signaling under fully defined conditions. Nature protocols 8, 162–175, doi:10.1038/nprot.2012.150 (2013). - DOI - PMC - PubMed
-
- Hashem SI et al. Brief Report: Oxidative Stress Mediates Cardiomyocyte Apoptosis in a Human Model of Danon Disease and Heart Failure. Stem cells 33, 2343–2350, doi:10.1002/stem.2015 (2015). - DOI - PMC - PubMed
-
- Shen Y et al. A map of the cis-regulatory sequences in the mouse genome. Nature 488, 116–120, doi:10.1038/nature11243 (2012). - DOI - PMC - PubMed
-
- Gorkin D et al. Systematic mapping of chromatin state landscapes during mouse development. bioRxiv (2017).
-
- Jolma A et al. DNA-binding specificities of human transcription factors. Cell 152, 327–339, doi:10.1016/j.cell.2012.12.009 (2013). - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
