

Emerging structural insights into glycosyltransferase-mediated synthesis of glycans
- PMID: 31427814
- PMCID: PMC6820136
- DOI: 10.1038/s41589-019-0350-2
Emerging structural insights into glycosyltransferase-mediated synthesis of glycans
Abstract
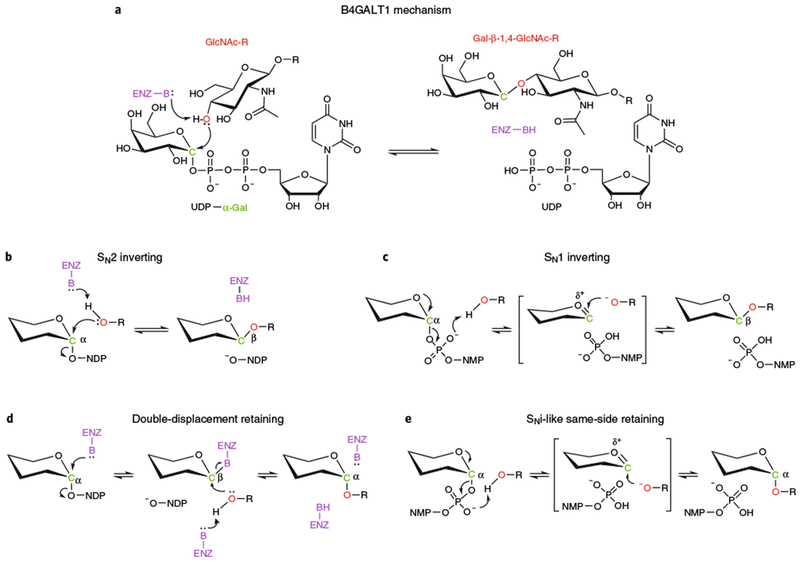
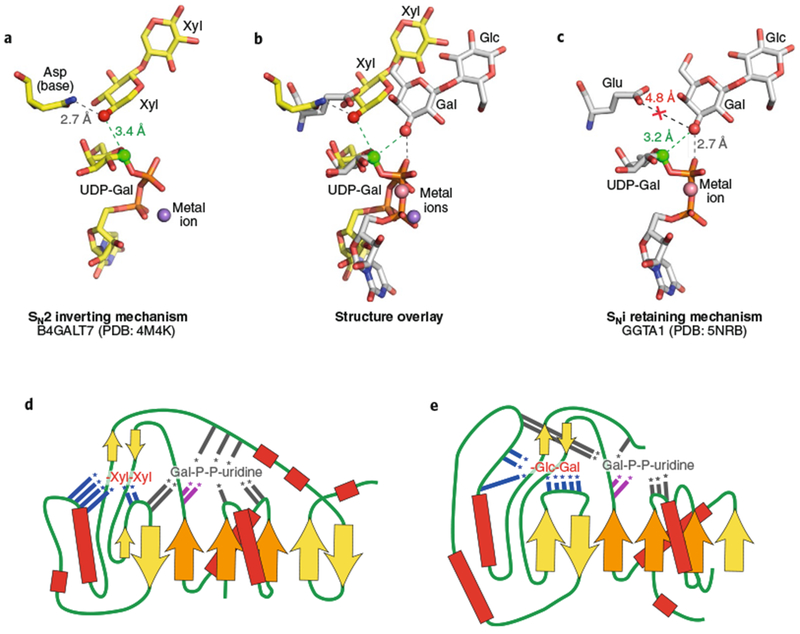
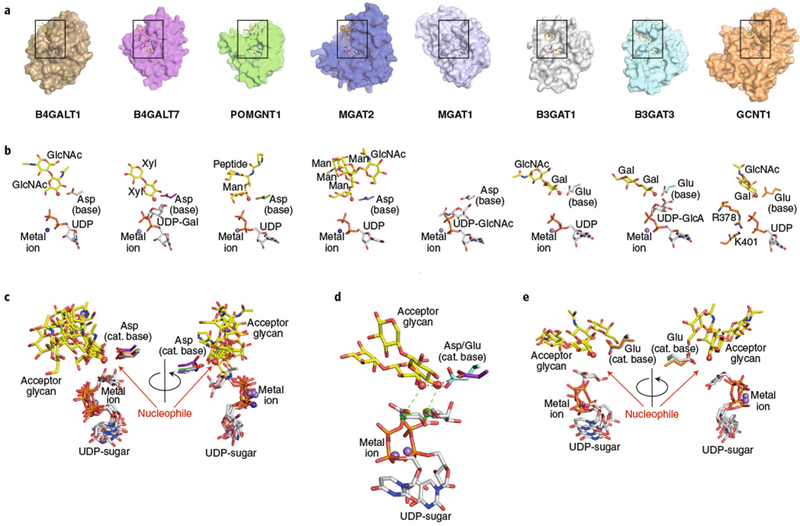
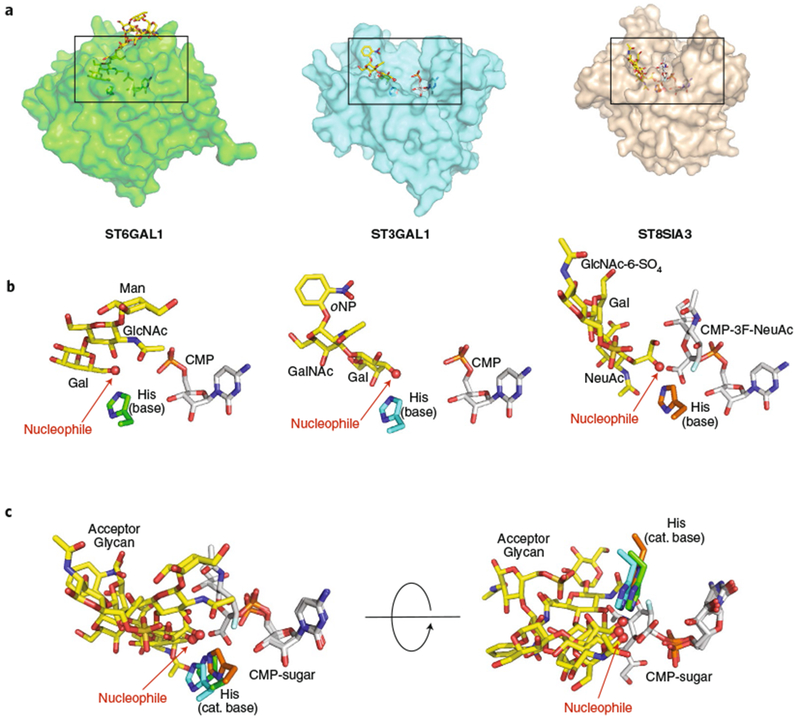
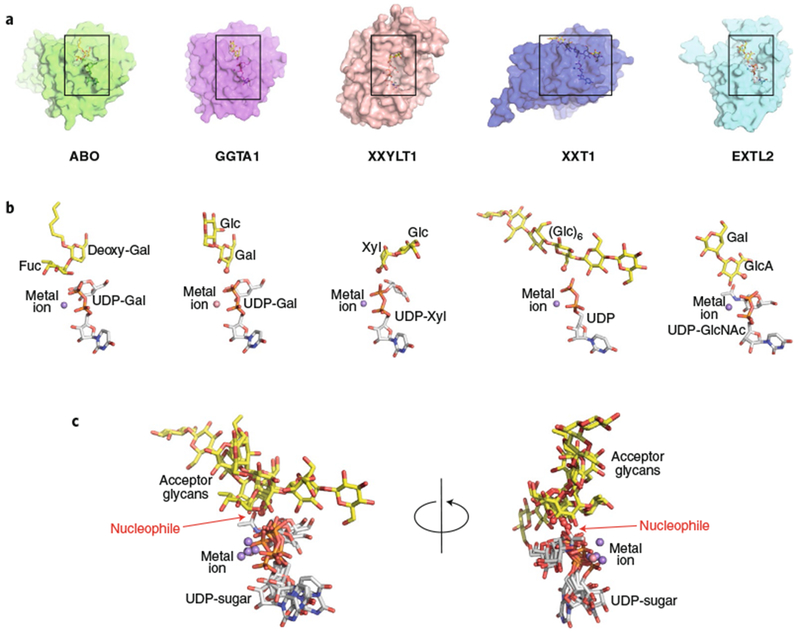
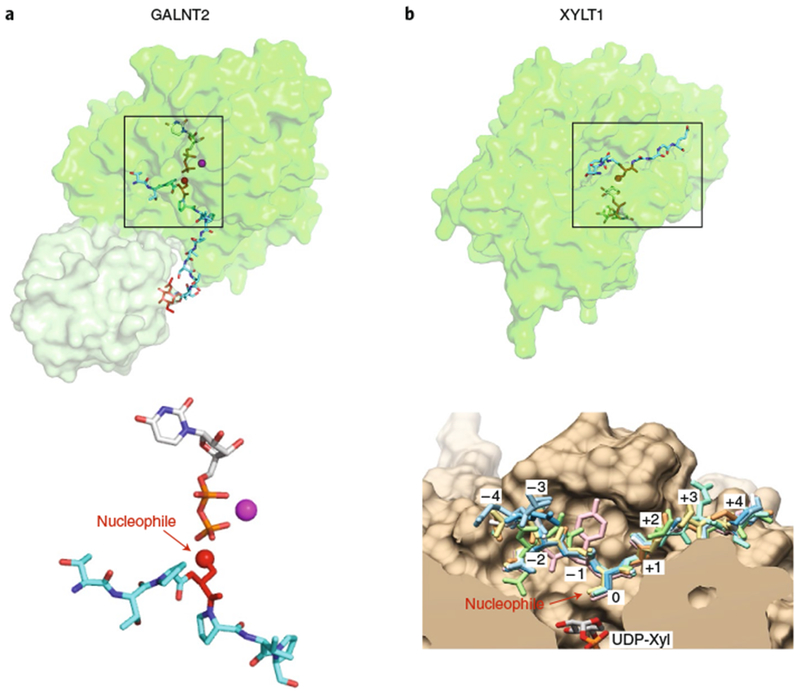
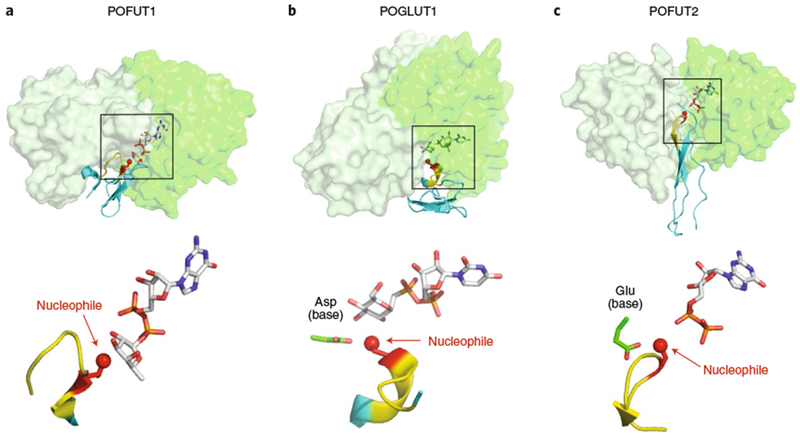
Glycans linked to proteins and lipids play key roles in biology; thus, accurate replication of cellular glycans is crucial for maintaining function following cell division. The fact that glycans are not copied from genomic templates suggests that fidelity is provided by the catalytic templates of glycosyltransferases that accurately add sugars to specific locations on growing oligosaccharides. To form new glycosidic bonds, glycosyltransferases bind acceptor substrates and orient a specific hydroxyl group, frequently one of many, for attack of the donor sugar anomeric carbon. Several recent crystal structures of glycosyltransferases with bound acceptor substrates reveal that these enzymes have common core structures that function as scaffolds upon which variable loops are inserted to confer substrate specificity and correctly orient the nucleophilic hydroxyl group. The varied approaches for acceptor binding site assembly suggest an ongoing evolution of these loop regions provides templates for assembly of the diverse glycan structures observed in biology.
Figures
References
-
- Lairson LL, Henrissat B, Davies GJ & Withers SG Glycosyltransferases: structures, functions, and mechanisms. Annu. Rev. Biochem 77, 521–555 (2008). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
