

Insect Odorscapes: From Plant Volatiles to Natural Olfactory Scenes
- PMID: 31427985
- PMCID: PMC6688386
- DOI: 10.3389/fphys.2019.00972
Insect Odorscapes: From Plant Volatiles to Natural Olfactory Scenes
Abstract
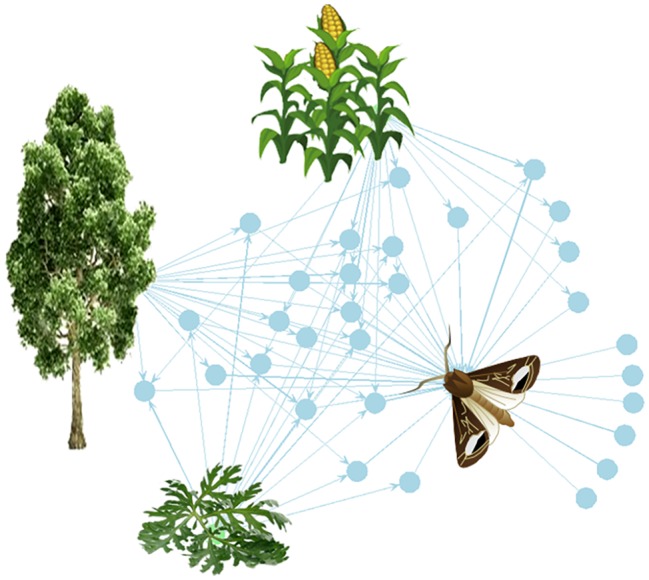
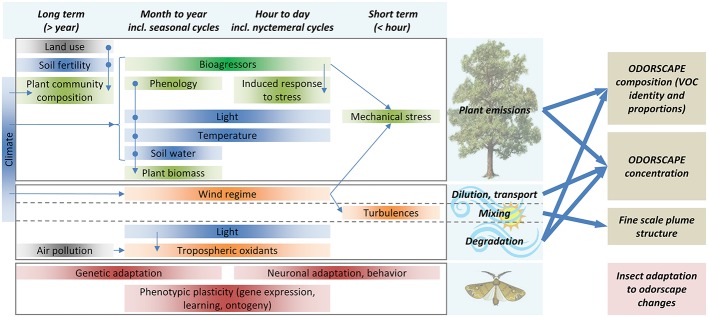
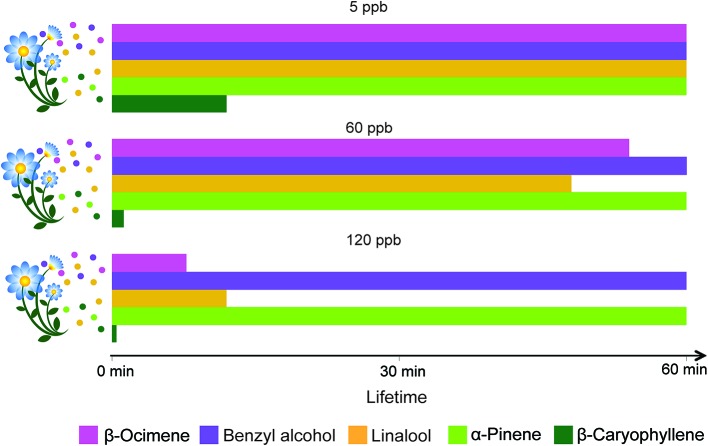
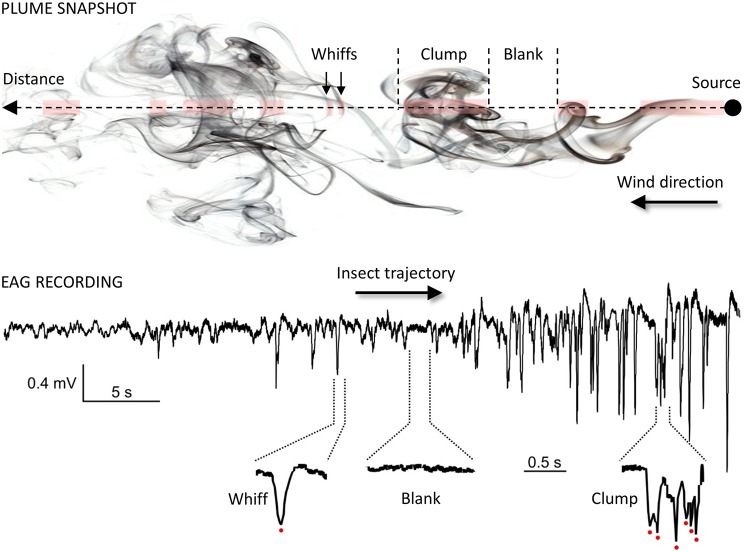
Olfaction is an essential sensory modality for insects and their olfactory environment is mostly made up of plant-emitted volatiles. The terrestrial vegetation produces an amazing diversity of volatile compounds, which are then transported, mixed, and degraded in the atmosphere. Each insect species expresses a set of olfactory receptors that bind part of the volatile compounds present in its habitat. Insect odorscapes are thus defined as species-specific olfactory spaces, dependent on the local habitat, and dynamic in time. Manipulations of pest-insect odorscapes are a promising approach to answer the strong demand for pesticide-free plant-protection strategies. Moreover, understanding their olfactory environment becomes a major concern in the context of global change and environmental stresses to insect populations. A considerable amount of information is available on the identity of volatiles mediating biotic interactions that involve insects. However, in the large body of research devoted to understanding how insects use olfaction to locate resources, an integrative vision of the olfactory environment has rarely been reached. This article aims to better apprehend the nature of the insect odorscape and its importance to insect behavioral ecology by reviewing the literature specific to different disciplines from plant ecophysiology to insect neuroethology. First, we discuss the determinants of odorscape composition, from the production of volatiles by plants (section "Plant Metabolism and Volatile Emissions") to their filtering during detection by the olfactory system of insects (section "Insect Olfaction: How Volatile Plant Compounds Are Encoded and Integrated by the Olfactory System"). We then summarize the physical and chemical processes by which volatile chemicals distribute in space (section "Transportation of Volatile Plant Compounds and Spatial Aspects of the Odorscape") and time (section "Temporal Aspects: The Dynamics of the Odorscape") in the atmosphere. The following sections consider the ecological importance of background odors in odorscapes and how insects adapt to their olfactory environment. Habitat provides an odor background and a sensory context that modulate the responses of insects to pheromones and other olfactory signals (section "Ecological Importance of Odorscapes"). In addition, insects do not respond inflexibly to single elements in their odorscape but integrate several components of their environment (section "Plasticity and Adaptation to Complex and Variable Odorscapes"). We finally discuss existing methods of odorscape manipulation for sustainable pest insect control and potential future developments in the context of agroecology (section "Odorscapes in Plant Protection and Agroecology").
Keywords: insect olfaction; landscape; odorscape; olfactome; plant volatiles; plant-insect interaction; sensory ecology; volatilome.
Figures
References
-
- Ache B. W., Gleeson R. A., Thompson H. A. (1988). Mechanisms for mixture suppression in olfactory receptors of the spiny lobster. Chem. Senses 13, 425–434. 10.1093/chemse/13.3.425 - DOI
-
- Andersson M. N. (2012). Mechanisms of odor coding in coniferous bark beetles: from neuron to behavior and application. Psyche. 2012:149572. 10.1155/2012/149572 - DOI
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources