

Monensin Sensitive 1 Regulates Dendritic Arborization in Drosophila by Modulating Endocytic Flux
- PMID: 31428611
- PMCID: PMC6687774
- DOI: 10.3389/fcell.2019.00145
Monensin Sensitive 1 Regulates Dendritic Arborization in Drosophila by Modulating Endocytic Flux
Abstract
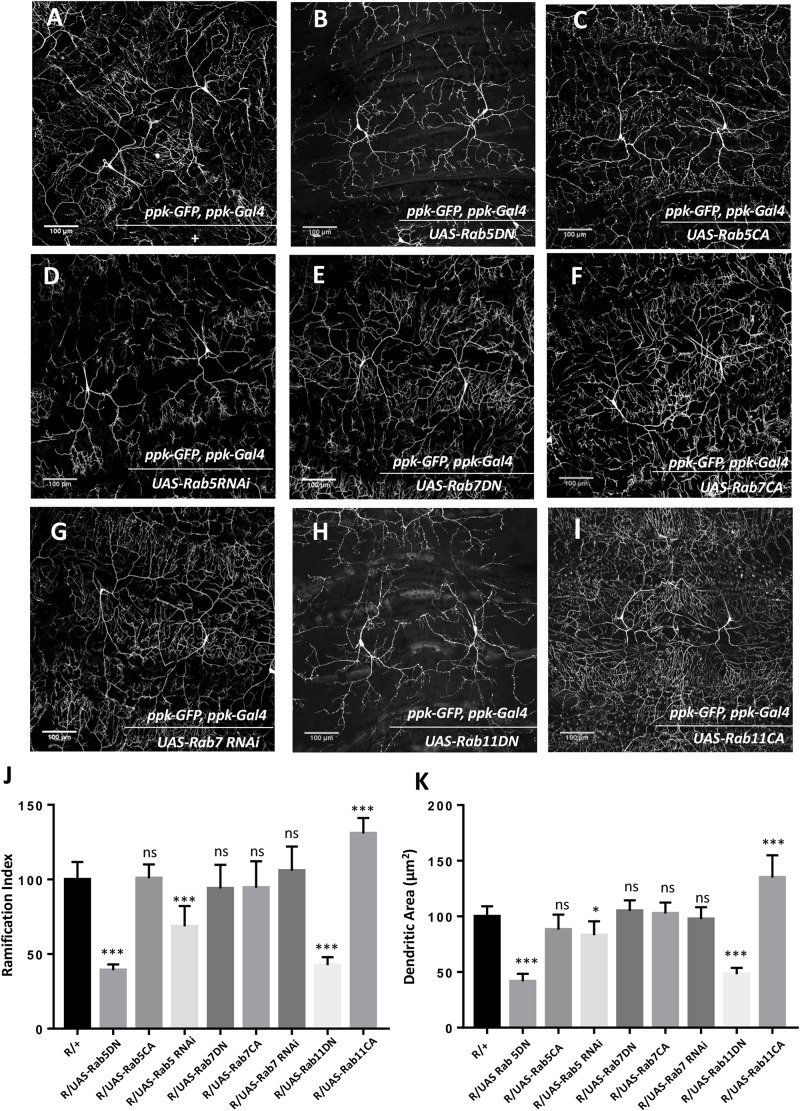
Monensin Sensitive 1 (Mon1) is a component of the Mon1:Ccz1 complex that mediates Rab5 to Rab7 conversion in eukaryotic cells by serving as a guanine nucleotide exchange factor for Rab7 during vesicular trafficking. We find that Mon1 activity modulates the complexity of Class IV dendritic arborization (da) neurons during larval development. Loss of Mon1 function leads to an increase in arborization and complexity, while increased expression, leads to reduced arborization. The ability of Mon1 to influence dendritic development is possibly a function of its interactions with Rab family GTPases that are central players in vesicular trafficking. Earlier, these GTPases, specifically Rab1, Rab5, Rab10, and Rab11 have been shown to regulate dendritic arborization. We have conducted genetic epistasis experiments, by modulating the activity of Rab5, Rab7, and Rab11 in da neurons, in Mon1 mutants, and demonstrate that the ability of Mon1 to regulate arborization is possibly due to its effect on the recycling pathway. Dendritic branching is critical for proper connectivity and physiological function of the neuron. An understanding of regulatory elements, such as Mon1, as demonstrated in our study, is essential to understand neuronal function.
Keywords: Class IV neuron; Rab conversion; endocytic recycling; epistasis; flux.
Figures



Similar articles
-
Activation of the Rab7 GTPase by the MON1-CCZ1 Complex Is Essential for PVC-to-Vacuole Trafficking and Plant Growth in Arabidopsis.Plant Cell. 2014 May;26(5):2080-2097. doi: 10.1105/tpc.114.123141. Epub 2014 May 13. Plant Cell. 2014. PMID: 24824487 Free PMC article.
-
Regulatory sites in the Mon1-Ccz1 complex control Rab5 to Rab7 transition and endosome maturation.Proc Natl Acad Sci U S A. 2023 Jul 25;120(30):e2303750120. doi: 10.1073/pnas.2303750120. Epub 2023 Jul 18. Proc Natl Acad Sci U S A. 2023. PMID: 37463208 Free PMC article.
-
A trimeric metazoan Rab7 GEF complex is crucial for endocytosis and scavenger function.J Cell Sci. 2020 Jul 9;133(13):jcs247080. doi: 10.1242/jcs.247080. J Cell Sci. 2020. PMID: 32499409
-
[Rab GTPases networks in membrane traffic in Saccharomyces cerevisiae].Yakugaku Zasshi. 2015;135(3):483-92. doi: 10.1248/yakushi.14-00246. Yakugaku Zasshi. 2015. PMID: 25759056 Review. Japanese.
-
Rab7: role of its protein interaction cascades in endo-lysosomal traffic.Cell Signal. 2011 Mar;23(3):516-21. doi: 10.1016/j.cellsig.2010.09.012. Epub 2010 Sep 21. Cell Signal. 2011. PMID: 20851765 Review.
References
-
- Brand A. H., Perrimon N. (1993). Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 118 401–415. - PubMed
-
- Cajal S. R. (1999). Texture of the Nervous System of Man and the Vertebrates. Berlin: Springer.
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous