

Growth and remodelling of living tissues: perspectives, challenges and opportunities
- PMID: 31431183
- PMCID: PMC6731508
- DOI: 10.1098/rsif.2019.0233
Growth and remodelling of living tissues: perspectives, challenges and opportunities
Abstract
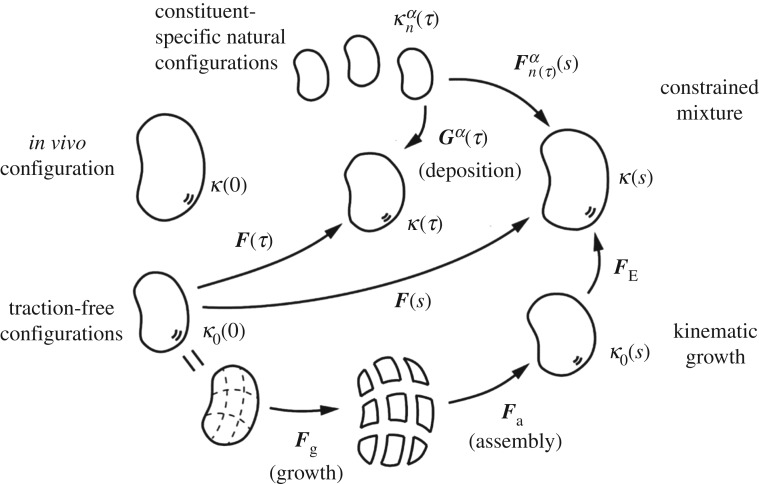
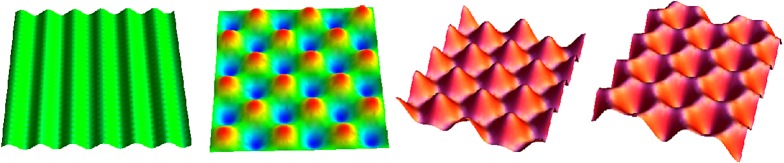
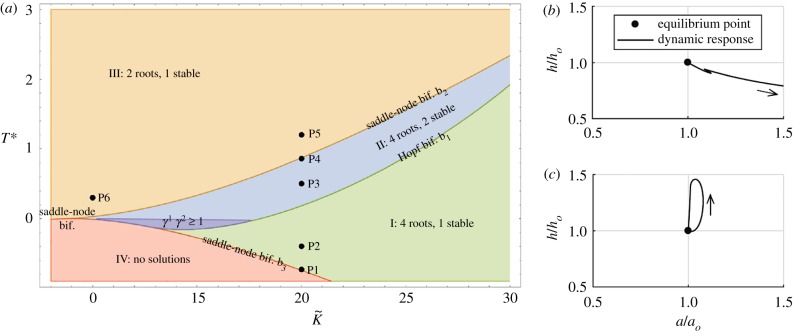
One of the most remarkable differences between classical engineering materials and living matter is the ability of the latter to grow and remodel in response to diverse stimuli. The mechanical behaviour of living matter is governed not only by an elastic or viscoelastic response to loading on short time scales up to several minutes, but also by often crucial growth and remodelling responses on time scales from hours to months. Phenomena of growth and remodelling play important roles, for example during morphogenesis in early life as well as in homeostasis and pathogenesis in adult tissues, which often adapt to changes in their chemo-mechanical environment as a result of ageing, diseases, injury or surgical intervention. Mechano-regulated growth and remodelling are observed in various soft tissues, ranging from tendons and arteries to the eye and brain, but also in bone, lower organisms and plants. Understanding and predicting growth and remodelling of living systems is one of the most important challenges in biomechanics and mechanobiology. This article reviews the current state of growth and remodelling as it applies primarily to soft tissues, and provides a perspective on critical challenges and future directions.
Keywords: growth; instabilities; living systems; morphoelasticity; remodelling.
Conflict of interest statement
We declare we have no competing interests.
Figures

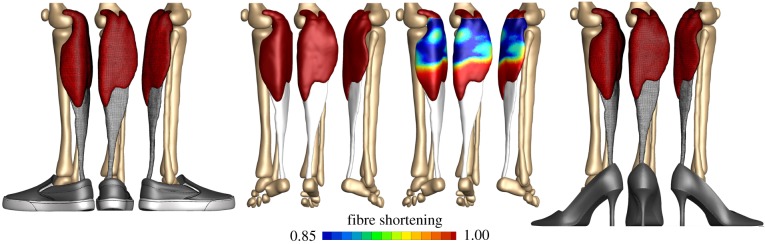
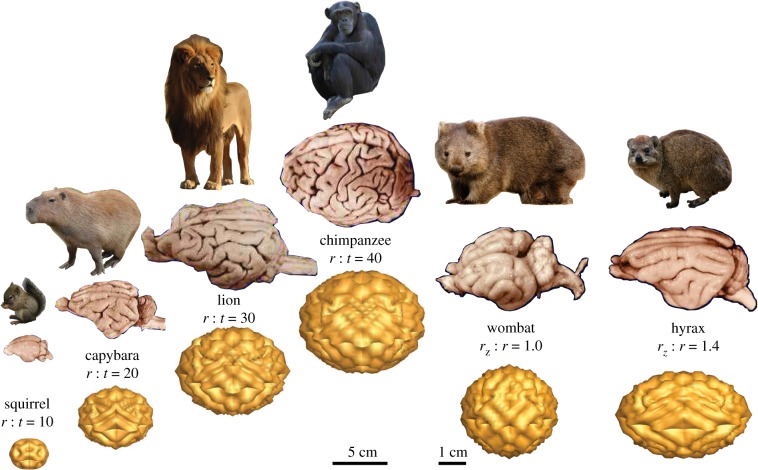
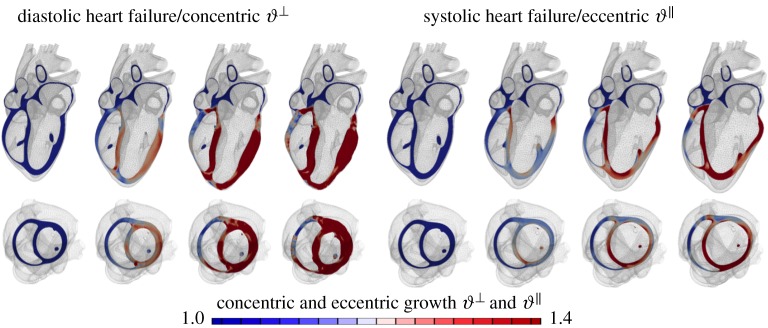
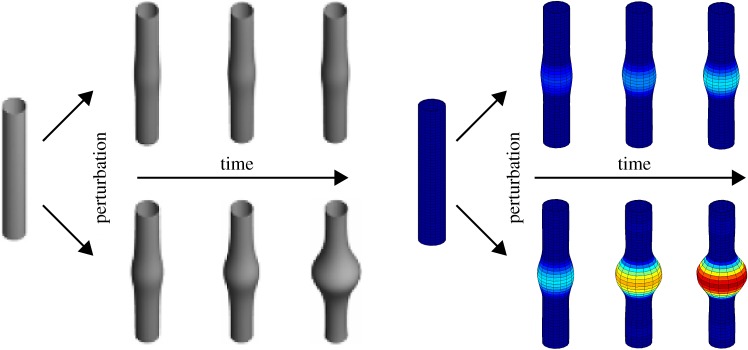
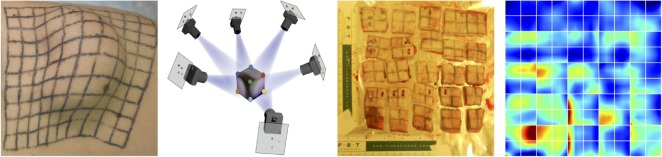
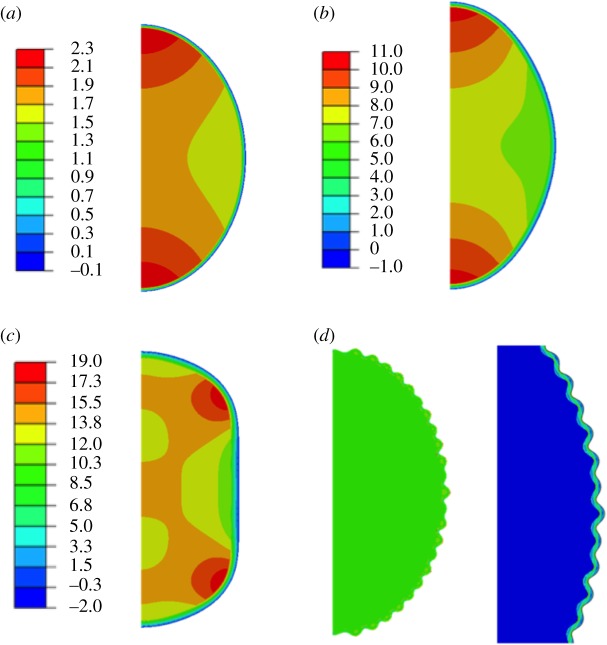
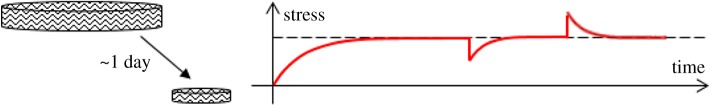
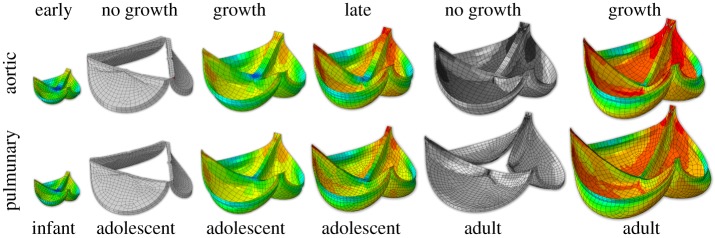
References
-
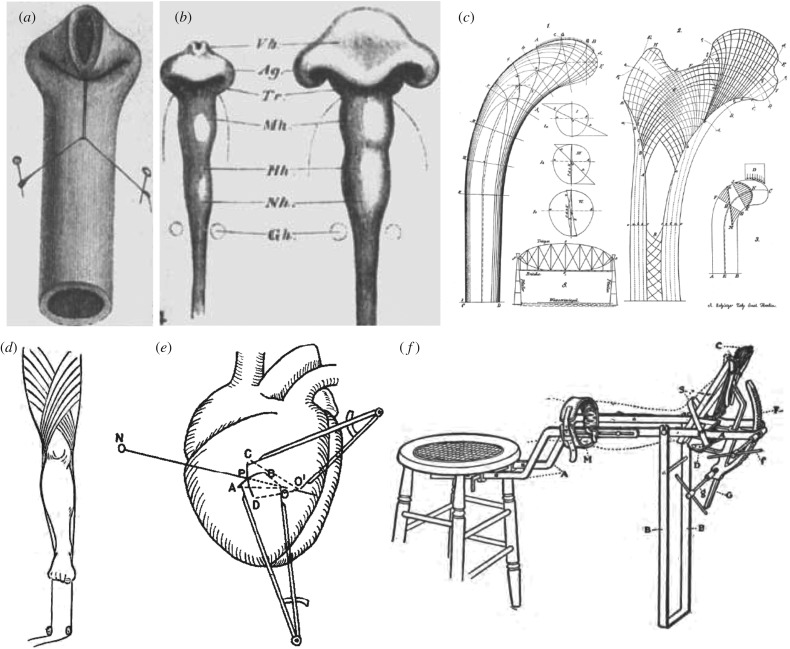
- His W. 1888. On the principles of animal morphology. Proc. R. Soc. Edin. 15, 287–298.
-
- Wolff J. 1870. Über die innere architektur der knochen und ihre bedeutung für die frage von knochenwachstum. Arch. Pathol. Anat. Physiol. Klin. Med. 50, 389–453. (10.1007/BF01944490) - DOI
-
- Davis HG. 1867. Conservative surgery. New York, NY: D. Appleton & Company.
-
- Nutt JJ. 1913. Diseases and deformities of the foot. London, UK: Forgotten Books.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
