

ADAMDEC1 Maintains a Growth Factor Signaling Loop in Cancer Stem Cells
- PMID: 31434712
- PMCID: PMC7400732
- DOI: 10.1158/2159-8290.CD-18-1308
ADAMDEC1 Maintains a Growth Factor Signaling Loop in Cancer Stem Cells
Abstract
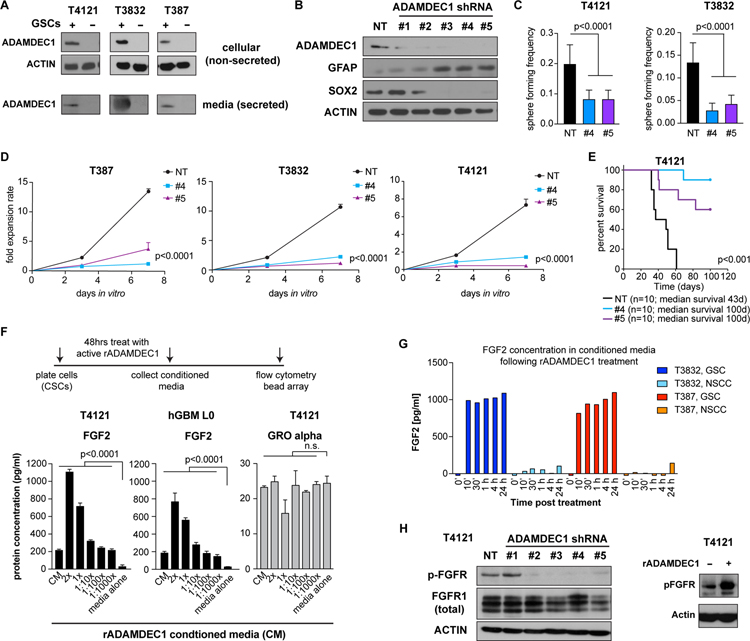
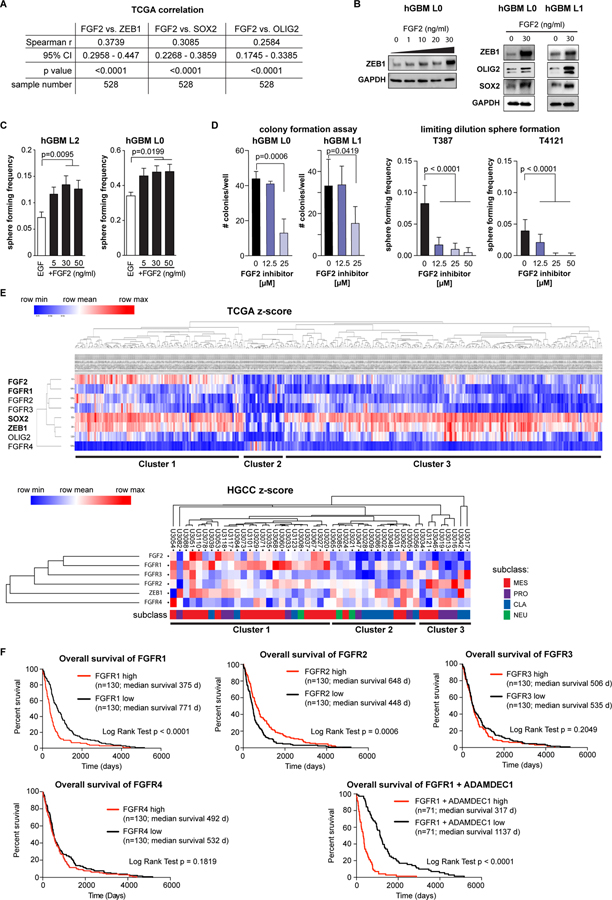
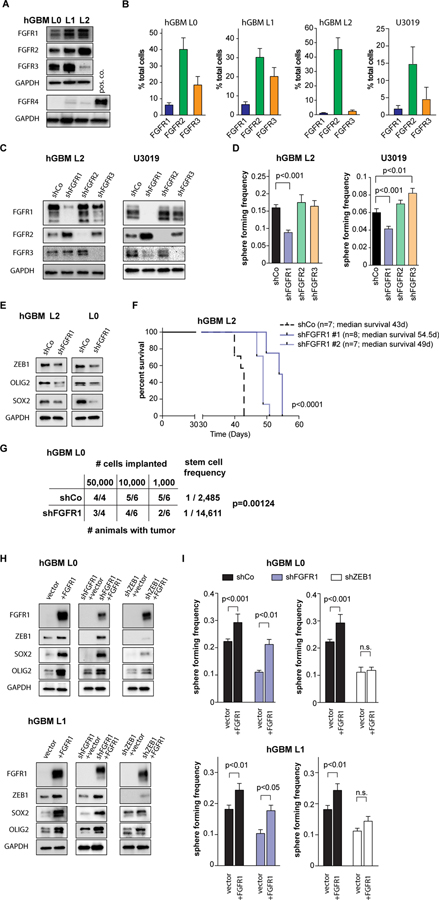
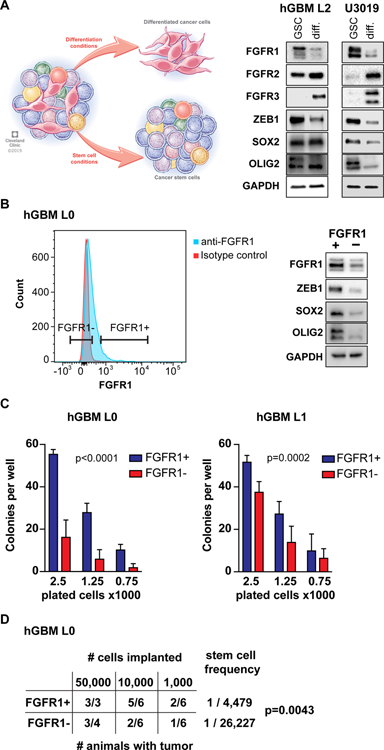
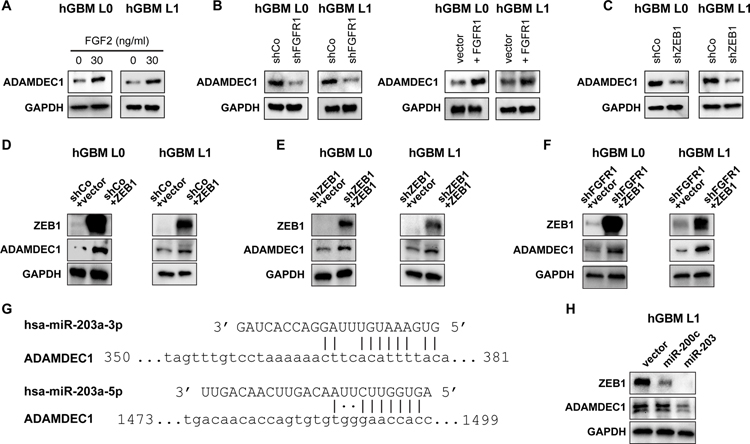
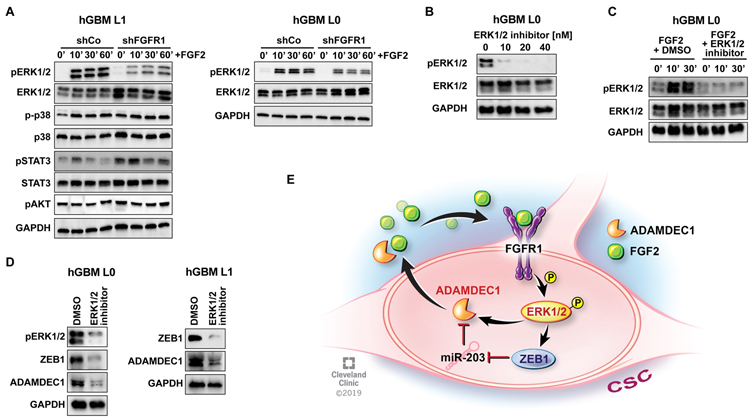
Glioblastomas (GBM) are lethal brain tumors where poor outcome is attributed to cellular heterogeneity, therapeutic resistance, and a highly infiltrative nature. These characteristics are preferentially linked to GBM cancer stem cells (GSC), but how GSCs maintain their stemness is incompletely understood and the subject of intense investigation. Here, we identify a novel signaling loop that induces and maintains GSCs consisting of an atypical metalloproteinase, ADAMDEC1, secreted by GSCs. ADAMDEC1 rapidly solubilizes FGF2 to stimulate FGFR1 expressed on GSCs. FGFR1 signaling induces upregulation of ZEB1 via ERK1/2 that regulates ADAMDEC1 expression through miR-203, creating a positive feedback loop. Genetic or pharmacologic targeting of components of this axis attenuates self-renewal and tumor growth. These findings reveal a new signaling axis for GSC maintenance and highlight ADAMDEC1 and FGFR1 as potential therapeutic targets in GBM. SIGNIFICANCE: Cancer stem cells (CSC) drive tumor growth in many cancers including GBM. We identified a novel sheddase, ADAMDEC1, which initiates an FGF autocrine loop to promote stemness in CSCs. This loop can be targeted to reduce GBM growth.This article is highlighted in the In This Issue feature, p. 1469.
©2019 American Association for Cancer Research.
Conflict of interest statement
Figures

Similar articles
-
Glutamine deprivation in glioblastoma stem cells triggers autophagic SIRT3 degradation to epigenetically restrict CD133 expression and stemness.Apoptosis. 2024 Oct;29(9-10):1619-1631. doi: 10.1007/s10495-024-02003-x. Epub 2024 Jul 28. Apoptosis. 2024. PMID: 39068621
-
METTL8 links mt-tRNA m3C modification to the HIF1α/RTK/Akt axis to sustain GBM stemness and tumorigenicity.Cell Death Dis. 2024 May 14;15(5):338. doi: 10.1038/s41419-024-06718-2. Cell Death Dis. 2024. PMID: 38744809 Free PMC article.
-
Glioblastoma stem cells deliver ABCB4 transcribed by ATF3 via exosomes conferring glioblastoma resistance to temozolomide.Cell Death Dis. 2024 May 6;15(5):318. doi: 10.1038/s41419-024-06695-6. Cell Death Dis. 2024. PMID: 38710703 Free PMC article.
-
Tumor heterogeneity and resistance in glioblastoma: the role of stem cells.Apoptosis. 2025 Aug;30(7-8):1695-1729. doi: 10.1007/s10495-025-02123-y. Epub 2025 May 15. Apoptosis. 2025. PMID: 40375039 Review.
-
Glioma Stem Cells as Promoter of Glioma Progression: A Systematic Review of Molecular Pathways and Targeted Therapies.Int J Mol Sci. 2024 Jul 22;25(14):7979. doi: 10.3390/ijms25147979. Int J Mol Sci. 2024. PMID: 39063221 Free PMC article.
Cited by
-
Small molecule-mediated disruption of ribosome biogenesis synergizes with FGFR inhibitors to suppress glioma cell growth.Neuro Oncol. 2023 Jun 2;25(6):1058-1072. doi: 10.1093/neuonc/noac286. Neuro Oncol. 2023. PMID: 36583853 Free PMC article.
-
Sex- and mutation-specific p53 gain-of-function activity in gliomagenesis.Cancer Res Commun. 2021 Dec;1(3):148-163. doi: 10.1158/2767-9764.crc-21-0026. Cancer Res Commun. 2021. PMID: 34957471 Free PMC article.
-
The Role of Exosomes in Thyroid Cancer and Their Potential Clinical Application.Front Oncol. 2020 Dec 1;10:596132. doi: 10.3389/fonc.2020.596132. eCollection 2020. Front Oncol. 2020. PMID: 33335859 Free PMC article. Review.
-
Multiorgan Signaling Mobilizes Tumor-Associated Erythroid Cells Expressing Immune Checkpoint Molecules.Mol Cancer Res. 2021 Mar;19(3):507-515. doi: 10.1158/1541-7786.MCR-20-0746. Epub 2020 Nov 24. Mol Cancer Res. 2021. PMID: 33234577 Free PMC article.
-
A transcriptome-wide association study integrating multi-omics bioinformatics and Mendelian randomization reveals the prognostic value of ADAMDEC1 in colon cancer.Arch Toxicol. 2025 Feb;99(2):645-665. doi: 10.1007/s00204-024-03910-3. Epub 2024 Dec 16. Arch Toxicol. 2025. PMID: 39680087
References
-
- Stupp R, Hegi ME, Mason WP, van den Bent MJ, Taphoorn MJ, Janzer RC, et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol 2009;10(5):459–66 10.1016/S1470-2045(09)70025-7. - DOI - PubMed
-
- Weller M, Butowski N, Tran DD, Recht LD, Lim M, Hirte H, et al. Rindopepimut with temozolomide for patients with newly diagnosed, EGFRvIII-expressing glioblastoma (ACT IV): a randomised, double-blind, international phase 3 trial. Lancet Oncol 2017;18(10):1373–85 10.1016/S1470-2045(17)30517-X. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
