

Wilm's tumor 1 promotes memory flexibility
- PMID: 31434897
- PMCID: PMC6704057
- DOI: 10.1038/s41467-019-11781-x
Wilm's tumor 1 promotes memory flexibility
Abstract
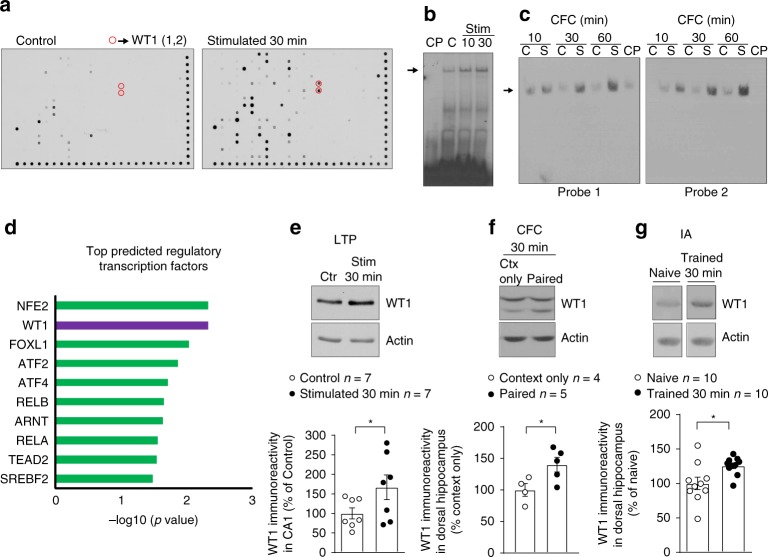
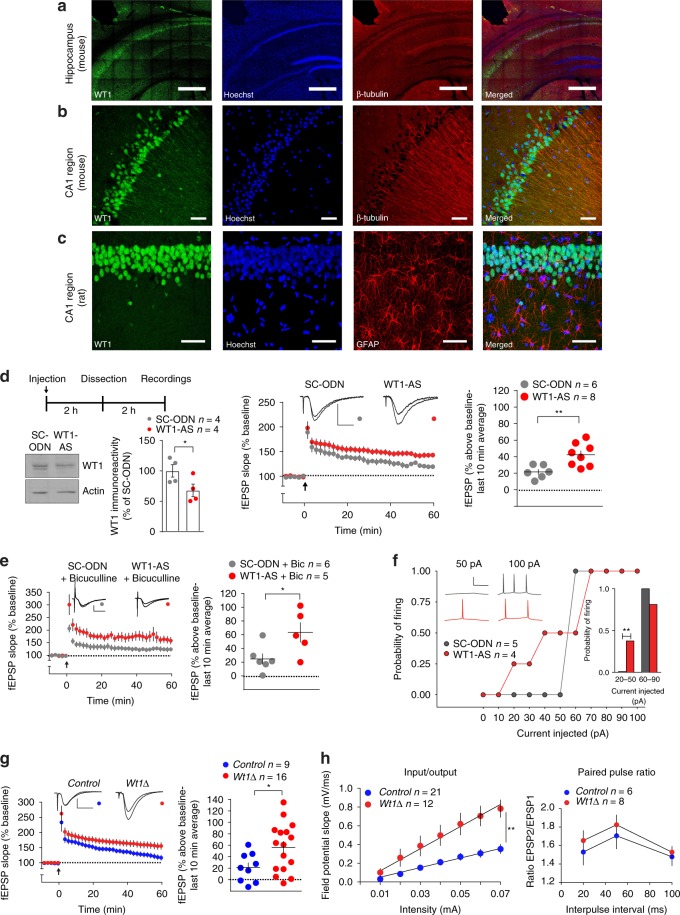
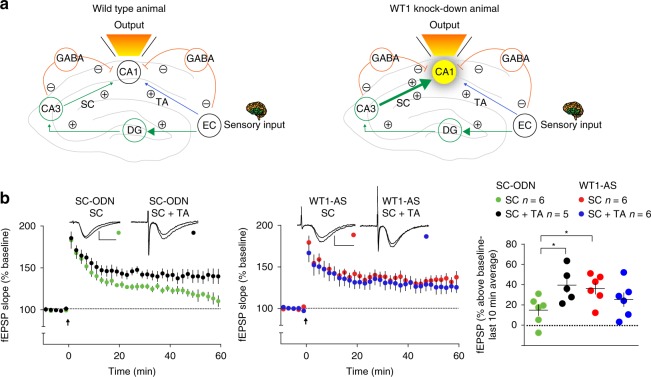
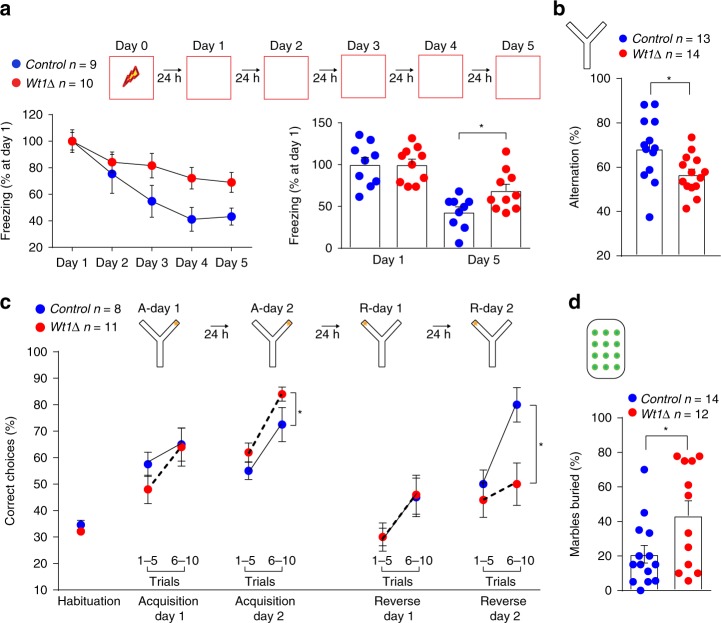
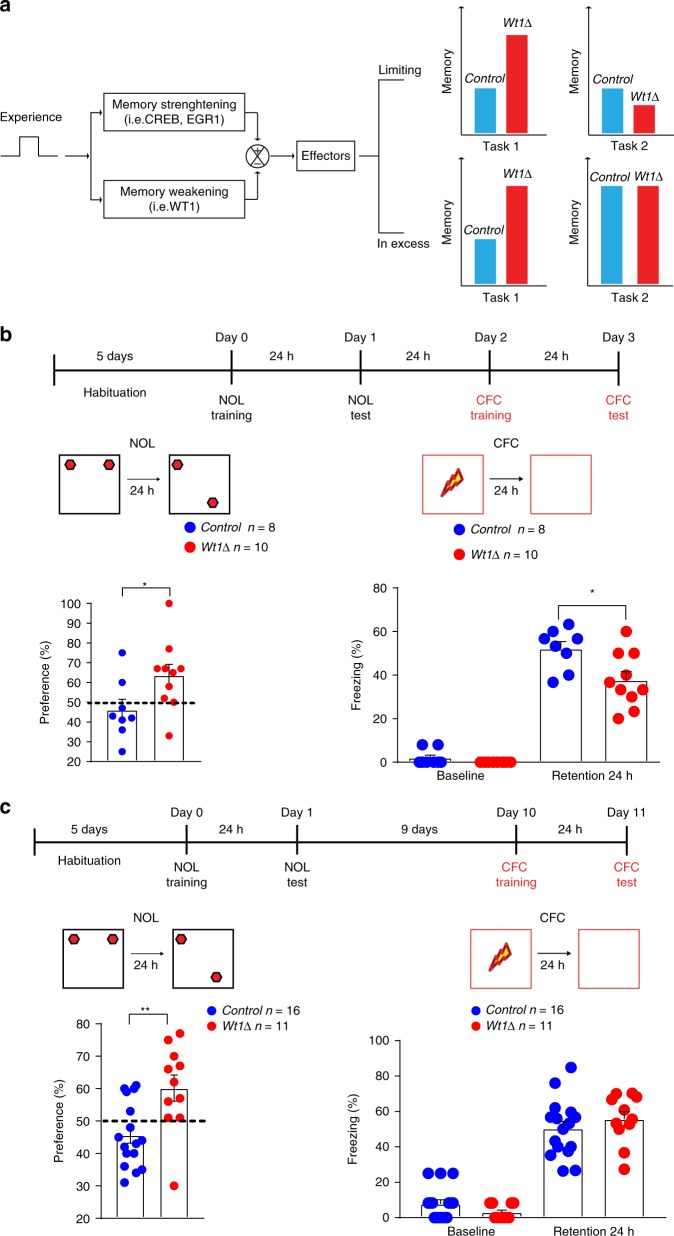
Under physiological conditions, strength and persistence of memory must be regulated in order to produce behavioral flexibility. In fact, impairments in memory flexibility are associated with pathologies such as post-traumatic stress disorder or autism; however, the underlying mechanisms that enable memory flexibility are still poorly understood. Here, we identify transcriptional repressor Wilm's Tumor 1 (WT1) as a critical synaptic plasticity regulator that decreases memory strength, promoting memory flexibility. WT1 is activated in the hippocampus following induction of long-term potentiation (LTP) or learning. WT1 knockdown enhances CA1 neuronal excitability, LTP and long-term memory whereas its overexpression weakens memory retention. Moreover, forebrain WT1-deficient mice show deficits in both reversal, sequential learning tasks and contextual fear extinction, exhibiting impaired memory flexibility. We conclude that WT1 limits memory strength or promotes memory weakening, thus enabling memory flexibility, a process that is critical for learning from new experiences.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
