

Conserved cell types with divergent features in human versus mouse cortex
- PMID: 31435019
- PMCID: PMC6919571
- DOI: 10.1038/s41586-019-1506-7
Conserved cell types with divergent features in human versus mouse cortex
Abstract
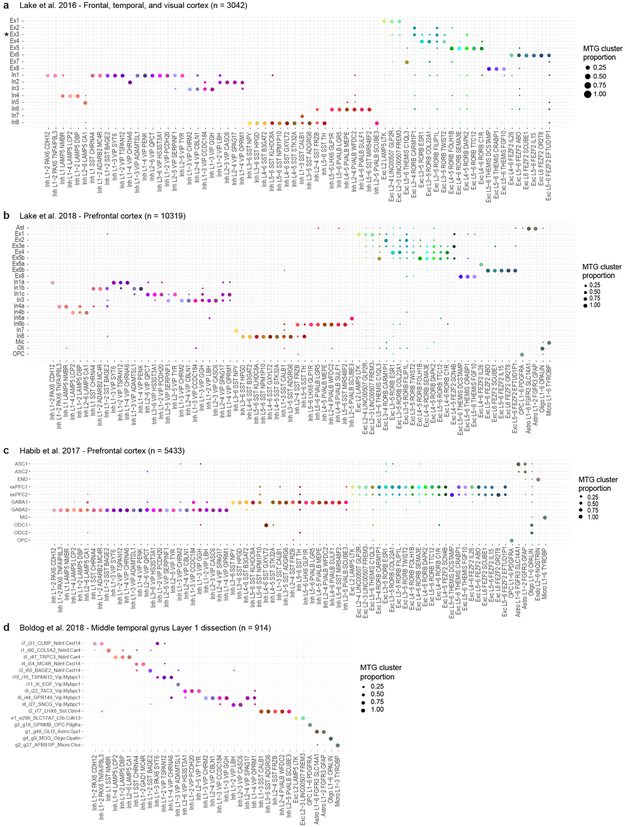
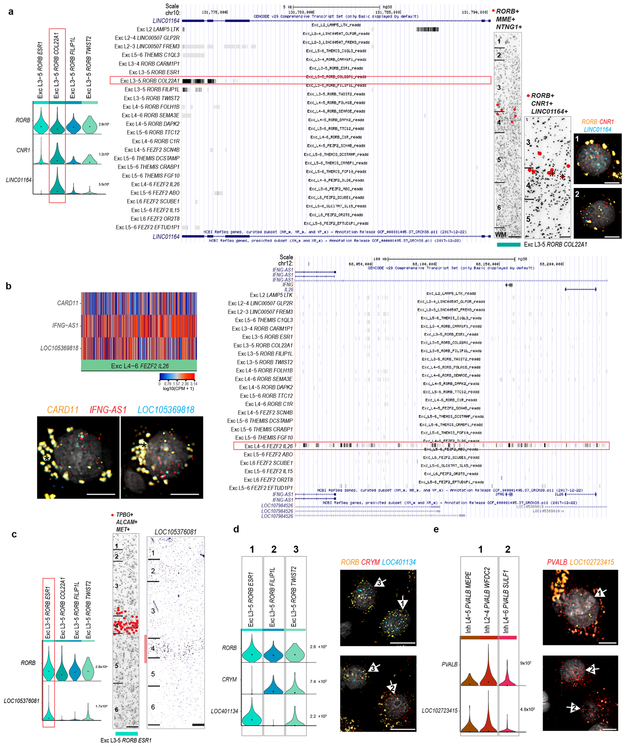
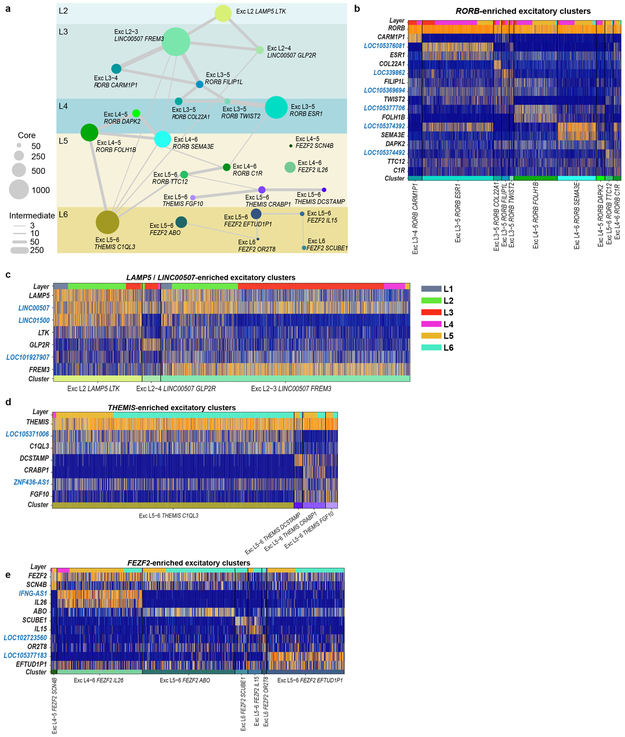
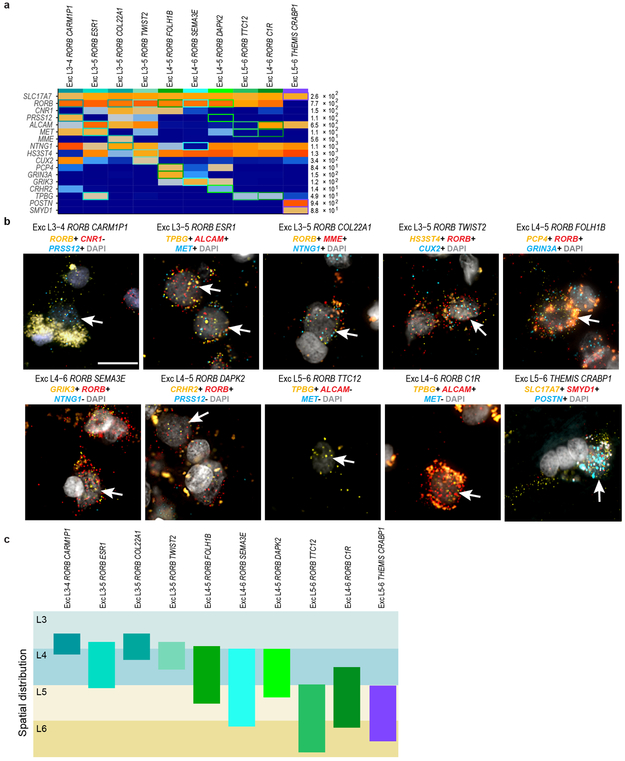
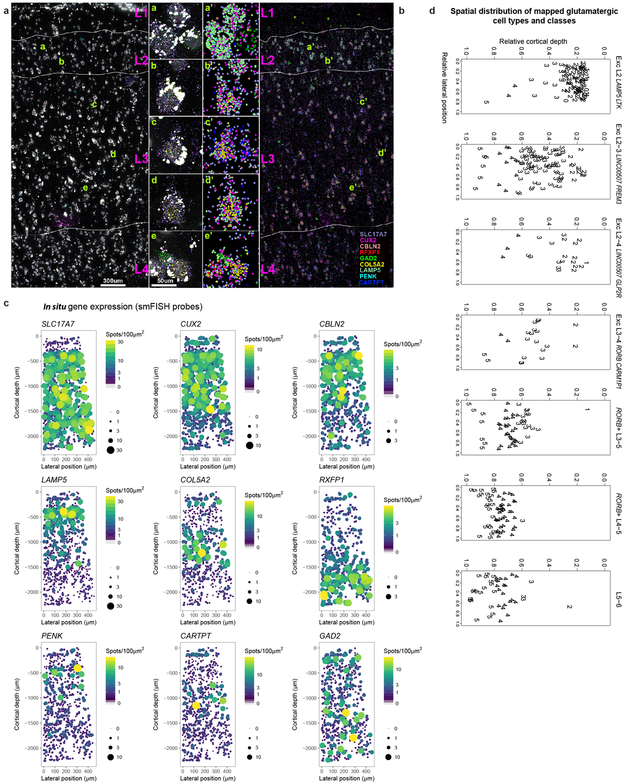
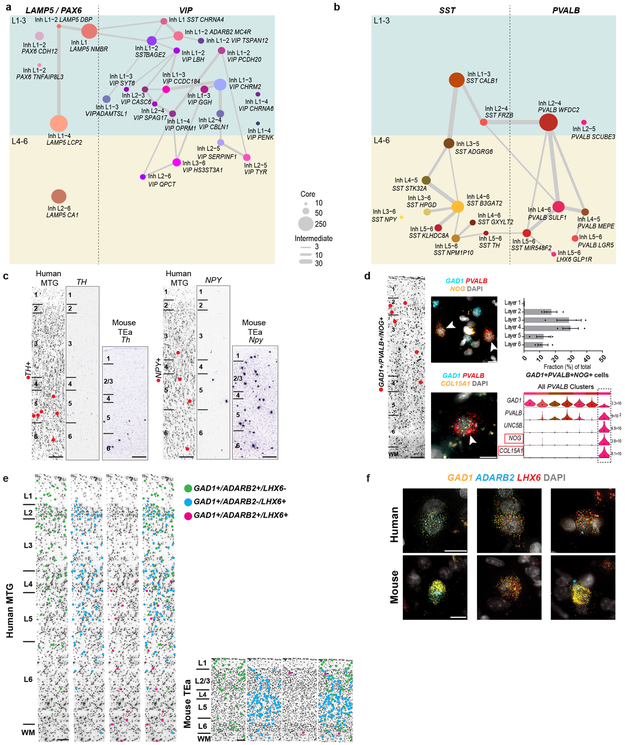
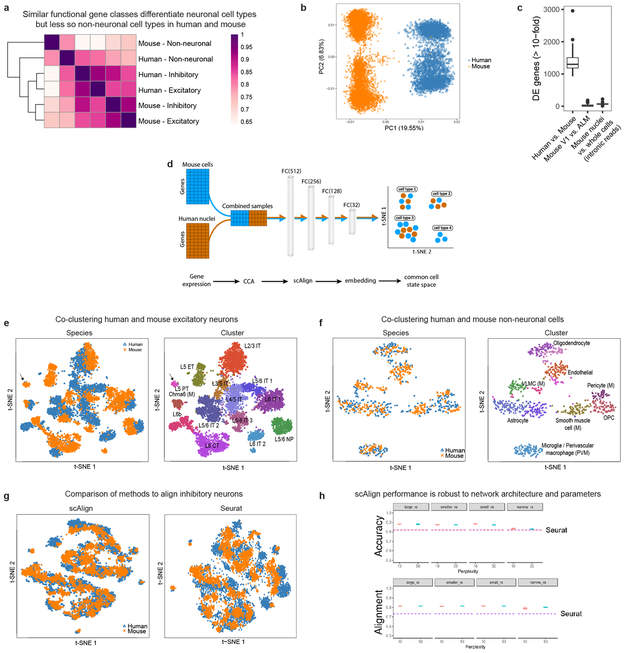
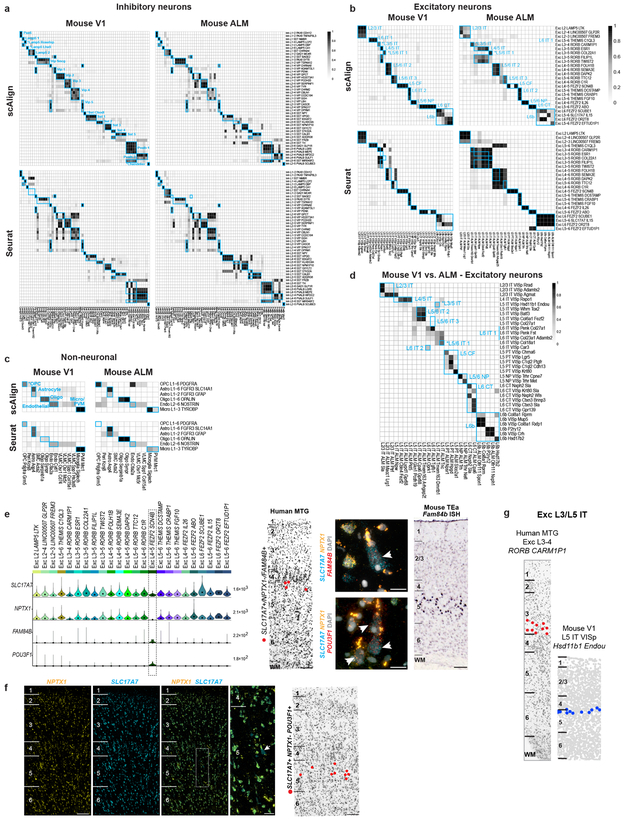
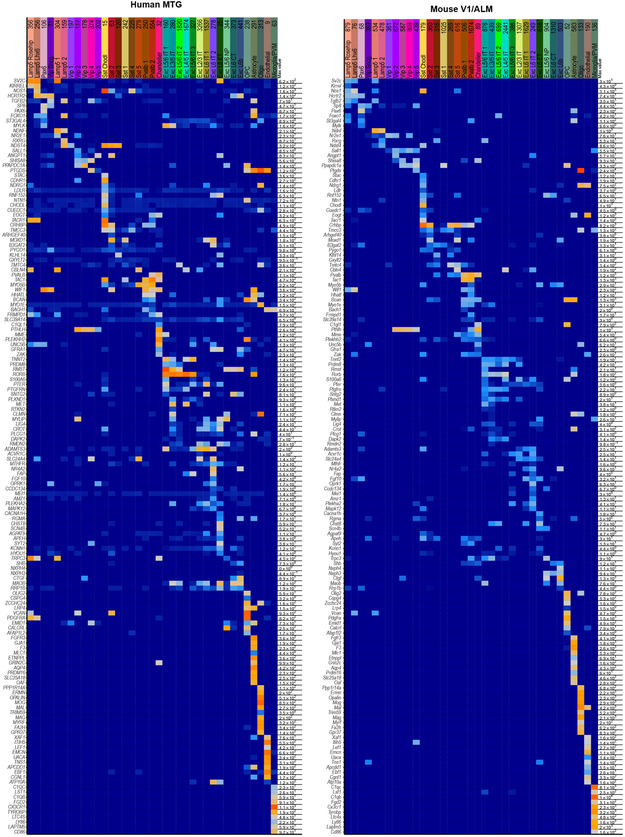
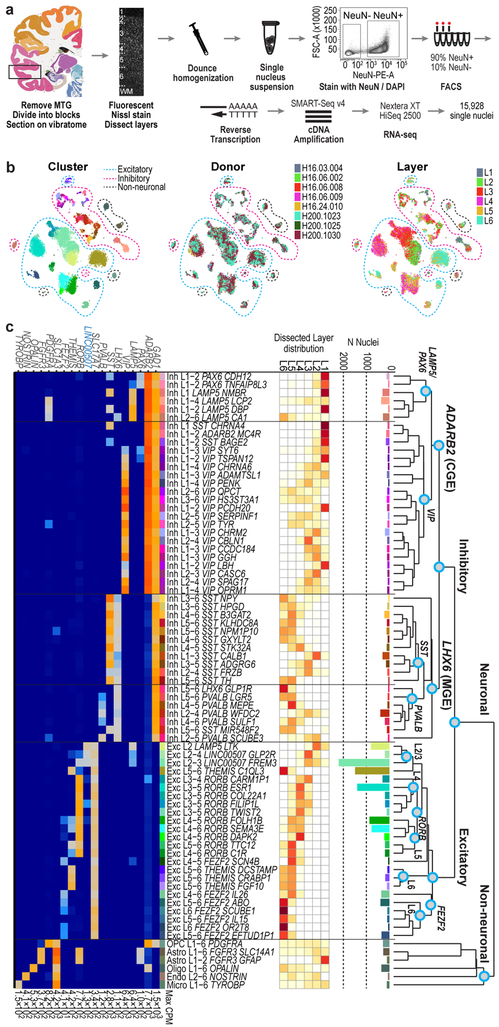
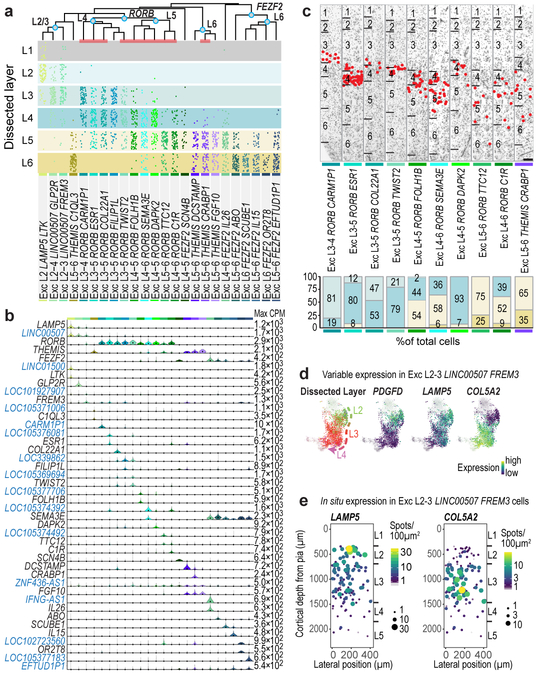
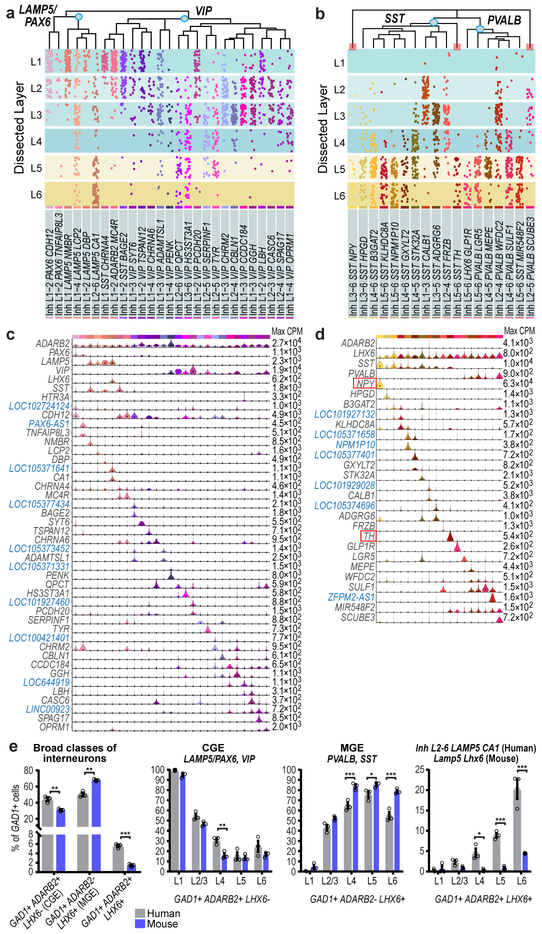
Elucidating the cellular architecture of the human cerebral cortex is central to understanding our cognitive abilities and susceptibility to disease. Here we used single-nucleus RNA-sequencing analysis to perform a comprehensive study of cell types in the middle temporal gyrus of human cortex. We identified a highly diverse set of excitatory and inhibitory neuron types that are mostly sparse, with excitatory types being less layer-restricted than expected. Comparison to similar mouse cortex single-cell RNA-sequencing datasets revealed a surprisingly well-conserved cellular architecture that enables matching of homologous types and predictions of properties of human cell types. Despite this general conservation, we also found extensive differences between homologous human and mouse cell types, including marked alterations in proportions, laminar distributions, gene expression and morphology. These species-specific features emphasize the importance of directly studying human brain.
Figures
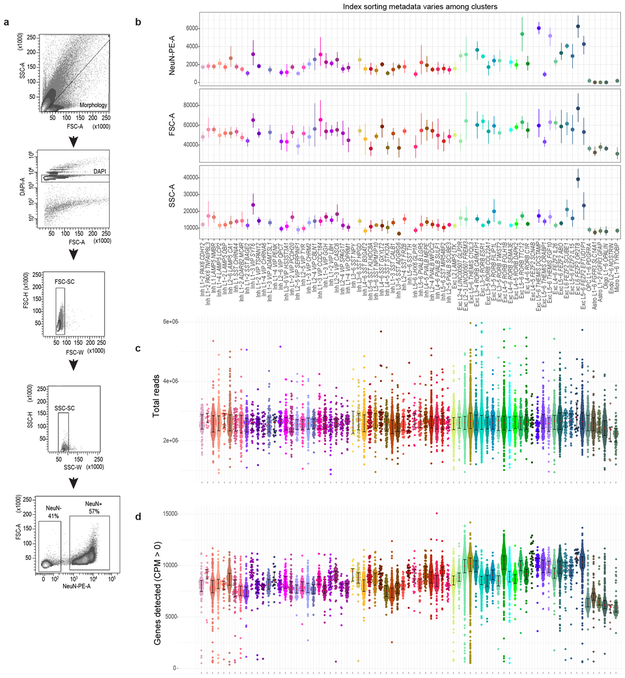
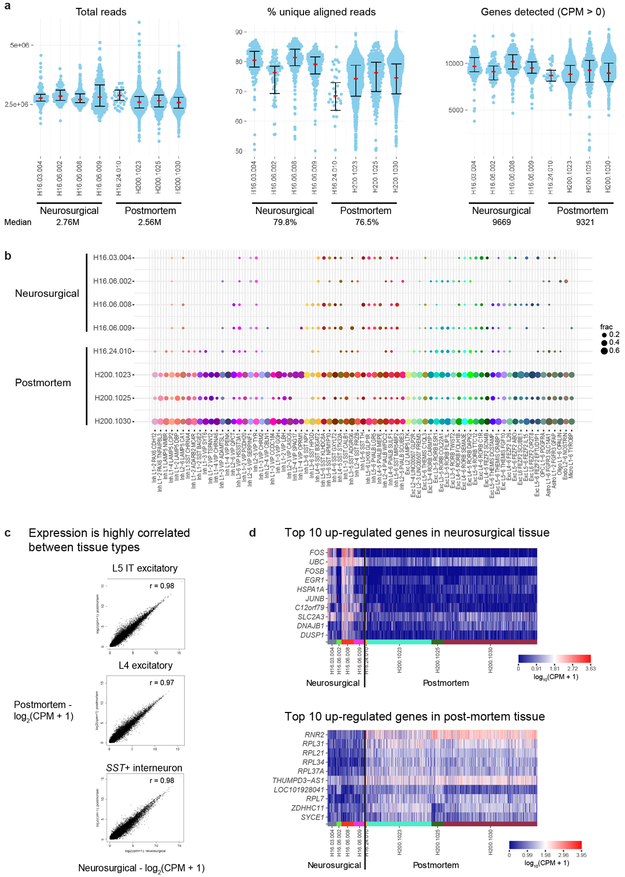
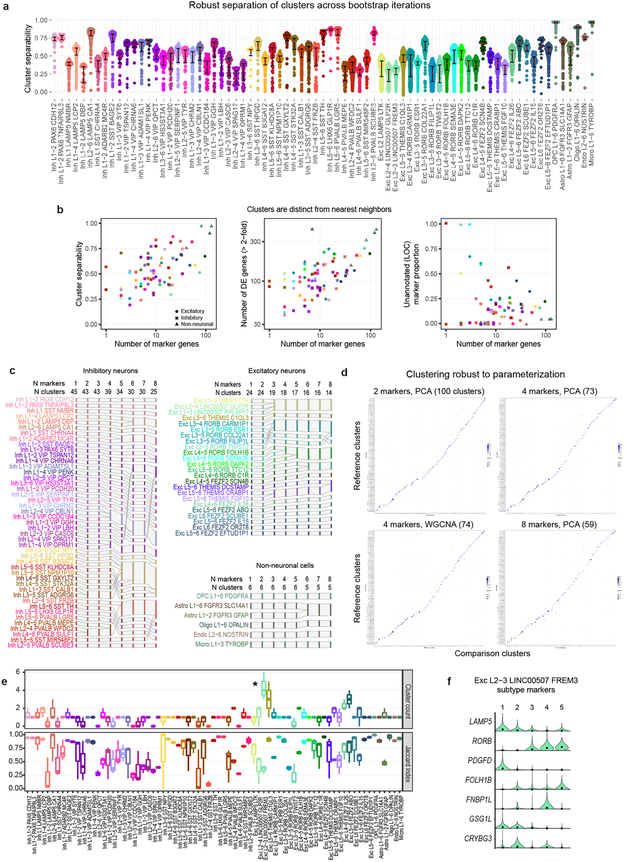
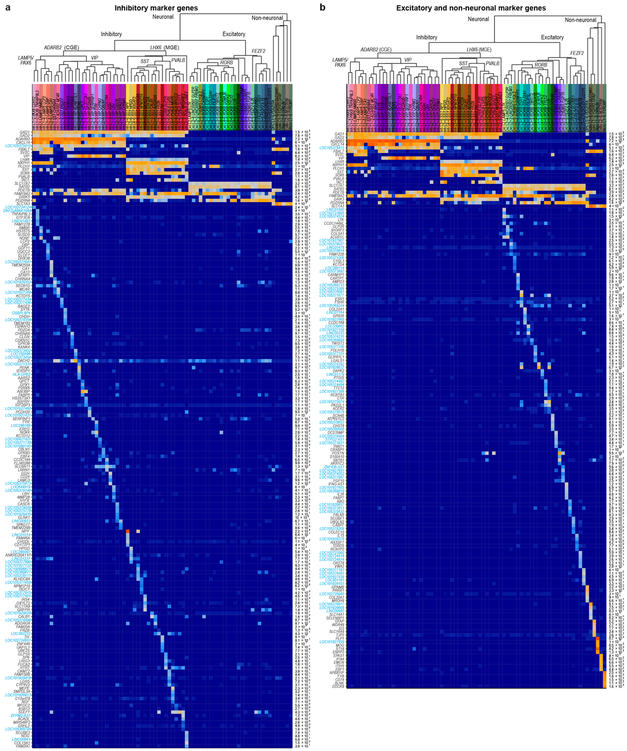
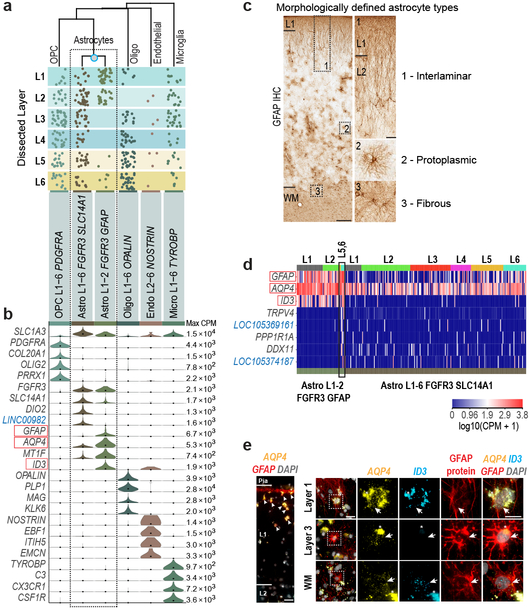
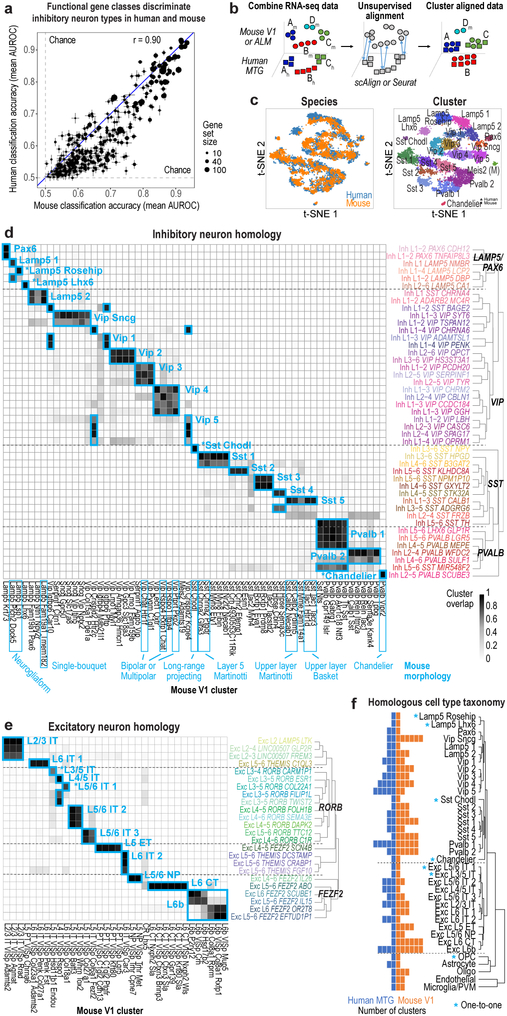
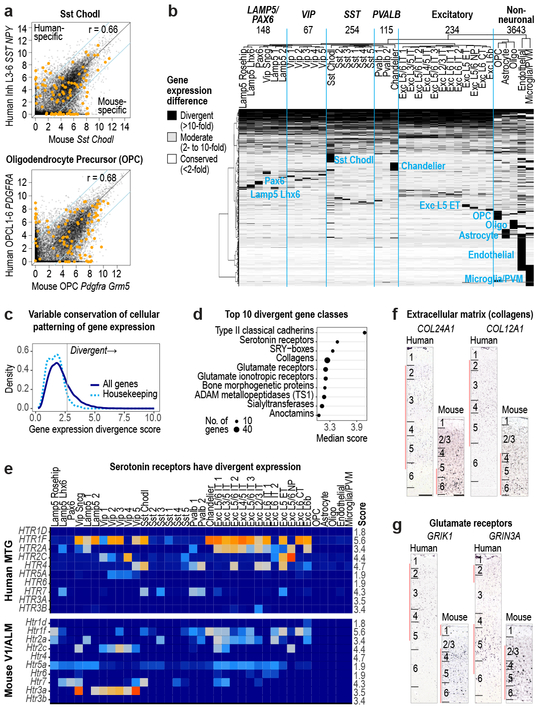
Comment in
-
A recipe book for cell types in the human brain.Nature. 2019 Sep;573(7772):36-37. doi: 10.1038/d41586-019-02343-8. Nature. 2019. PMID: 31481767 No abstract available.
-
Delving into diversity.Nat Rev Neurosci. 2019 Nov;20(11):646-647. doi: 10.1038/s41583-019-0228-z. Nat Rev Neurosci. 2019. PMID: 31548715 No abstract available.
-
Species-specific cortex features.Nat Cell Biol. 2021 Nov;23(11):1108. doi: 10.1038/s41556-021-00777-9. Nat Cell Biol. 2021. PMID: 34750576 No abstract available.
References
-
- Azevedo FAC et al. Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. The Journal of Comparative Neurology 513, 532–541 (2009). - PubMed
Methods References
-
- DeFelipe J, Alonso-Nanclares L & Arellano JI Microstructure of the neocortex: comparative aspects. J Neurocytol 31, 299–316 (2002). - PubMed
-
- Aronesty E Comparison of sequencing utility programs. The Open Bioinformatics Journal 7, 1–8 (2013).
