

SIRT6, a Mammalian Deacylase with Multitasking Abilities
- PMID: 31437090
- PMCID: PMC7002868
- DOI: 10.1152/physrev.00030.2018
SIRT6, a Mammalian Deacylase with Multitasking Abilities
Abstract
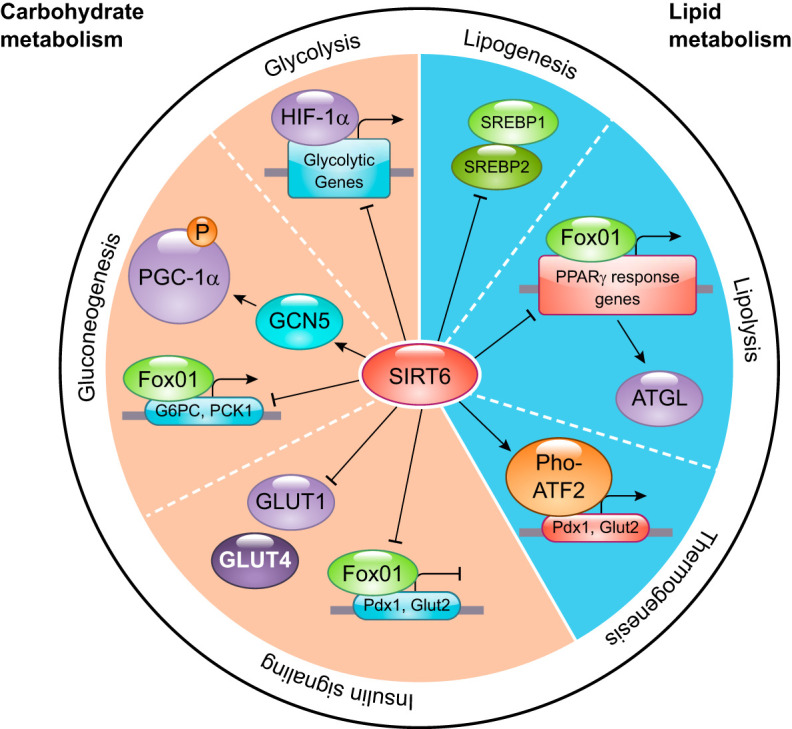
Mammalian sirtuins have emerged in recent years as critical modulators of multiple biological processes, regulating cellular metabolism, DNA repair, gene expression, and mitochondrial biology. As such, they evolved to play key roles in organismal homeostasis, and defects in these proteins have been linked to a plethora of diseases, including cancer, neurodegeneration, and aging. In this review, we describe the multiple roles of SIRT6, a chromatin deacylase with unique and important functions in maintaining cellular homeostasis. We attempt to provide a framework for such different functions, for the ability of SIRT6 to interconnect chromatin dynamics with metabolism and DNA repair, and the open questions the field will face in the future, particularly in the context of putative therapeutic opportunities.
Keywords: SIRT6; cancer; chromatin deacetylase; epigenetics.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures






References
-
- Abifadel M, Varret M, Rabès JP, Allard D, Ouguerram K, Devillers M, Cruaud C, Benjannet S, Wickham L, Erlich D, Derré A, Villéger L, Farnier M, Beucler I, Bruckert E, Chambaz J, Chanu B, Lecerf JM, Luc G, Moulin P, Weissenbach J, Prat A, Krempf M, Junien C, Seidah NG, Boileau C. Mutations in PCSK9 cause autosomal dominant hypercholesterolemia. Nat Genet 34: 154–156, 2003. doi: 10.1038/ng1161. - DOI - PubMed
-
- Atsumi Y, Minakawa Y, Ono M, Dobashi S, Shinohe K, Shinohara A, Takeda S, Takagi M, Takamatsu N, Nakagama H, Teraoka H, Yoshioka K. ATM and SIRT6/SNF2H Mediate Transient H2AX Stabilization When DSBs Form by Blocking HUWE1 to Allow Efficient γH2AX Foci Formation. Cell Rep 13: 2728–2740, 2015. doi: 10.1016/j.celrep.2015.11.054. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials