

Differential tetraspanin genes expression and subcellular localization during mutualistic interactions in Phaseolus vulgaris
- PMID: 31437164
- PMCID: PMC6705802
- DOI: 10.1371/journal.pone.0219765
Differential tetraspanin genes expression and subcellular localization during mutualistic interactions in Phaseolus vulgaris
Abstract
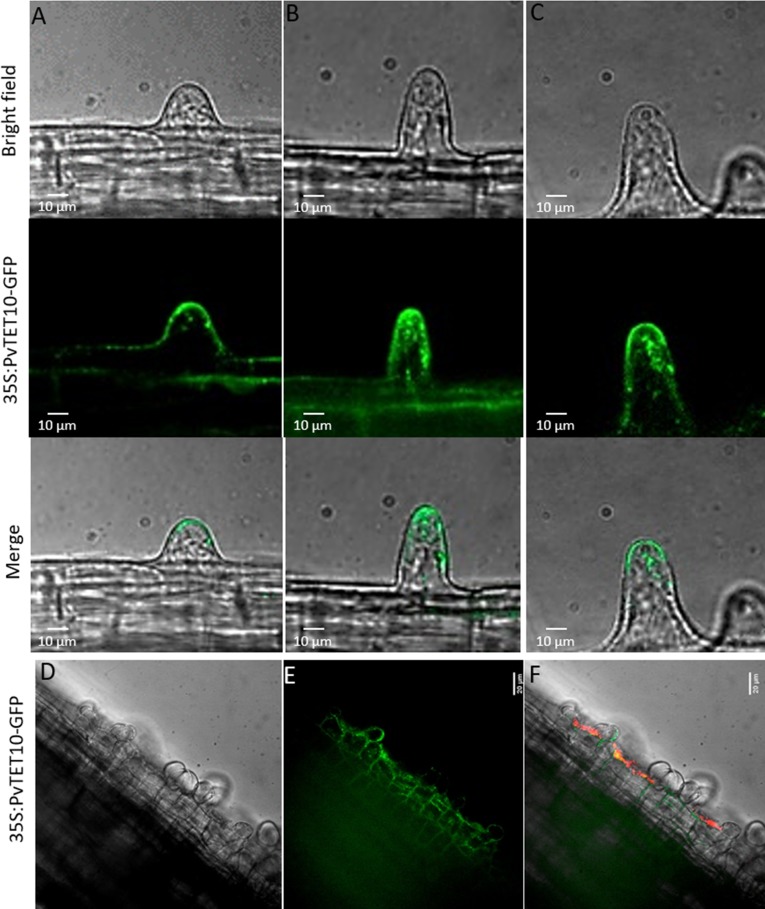
Arbuscular mycorrhizal fungi and rhizobia association with plants are two of the most successful plant-microbe associations that allow the assimilation of P and N by plants, respectively. These mutualistic interactions require a molecular dialogue, i.e., legume roots exude flavonoids or strigolactones which induce the Nod factors or Myc factors synthesis and secretion from the rhizobia or fungi, respectively. These Nod or Myc factors trigger several responses in the plant root, including calcium oscillations, and reactive oxygen species (ROS). Furthermore, superoxide and H2O2 have emerged as key components that regulate the transitions from proliferation to differentiation in the plant meristems. Similar to the root meristem, the nodule meristem accumulates superoxide and H2O2. Tetraspanins are transmembrane proteins that organize into tetraspanin web regions, where they recruit specific proteins into platforms required for signal transduction, membrane fusion, cell trafficking and ROS generation. Plant tetraspanins are scaffolding proteins associated with root radial patterning, biotic and abiotic stress responses, cell fate determination, and hormonal regulation and recently have been reported as a specific marker of exosomes in animal and plant cells and key players at the site of plant fungal infection. In this study, we conducted transcriptional profiling of the tetraspanin family in common bean (Phaseolus vulgaris L. var. Negro Jamapa) to determine the specific expression patterns and subcellular localization of tetraspanins during nodulation or under mycorrhizal association. Our results demonstrate that the tetraspanins are transcriptionally modulated during the mycorrhizal association, but are also expressed in the infection thread and nodule meristem development. Subcellular localization indicates that tetraspanins have a key role in vesicular trafficking, cell division, and root hair polar growth.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures










References
-
- Zepeda I, Sanchez-Lopez R, Kunkel JG, Banuelos LA, Hernandez-Barrera A, Sanchez F, et al. Visualization of highly dynamic F-actin plus ends in growing phaseolus vulgaris root hair cells and their responses to Rhizobium etli nod factors. Plant & cell physiology. 2014;55(3):580–92. 10.1093/pcp/pct202 . - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
