

Base Editor Correction of COL7A1 in Recessive Dystrophic Epidermolysis Bullosa Patient-Derived Fibroblasts and iPSCs
- PMID: 31437443
- PMCID: PMC6983342
- DOI: 10.1016/j.jid.2019.07.701
Base Editor Correction of COL7A1 in Recessive Dystrophic Epidermolysis Bullosa Patient-Derived Fibroblasts and iPSCs
Abstract
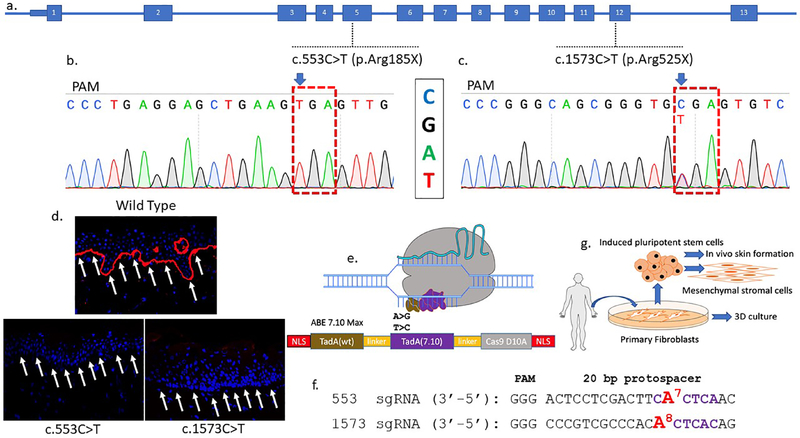
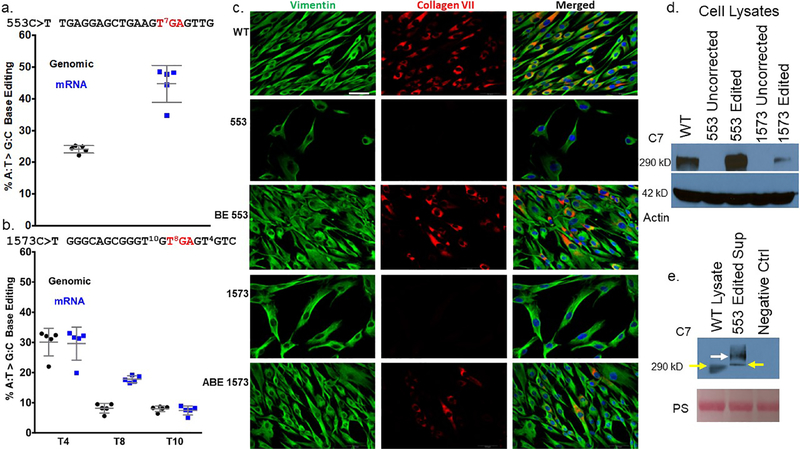
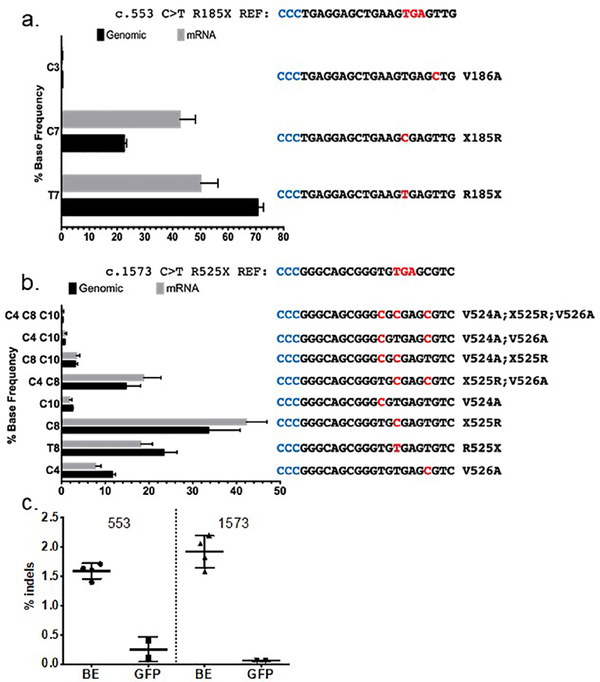
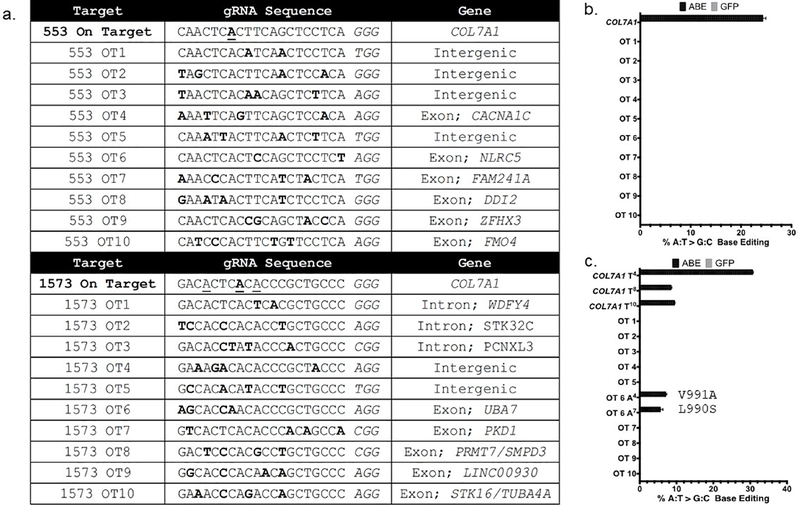
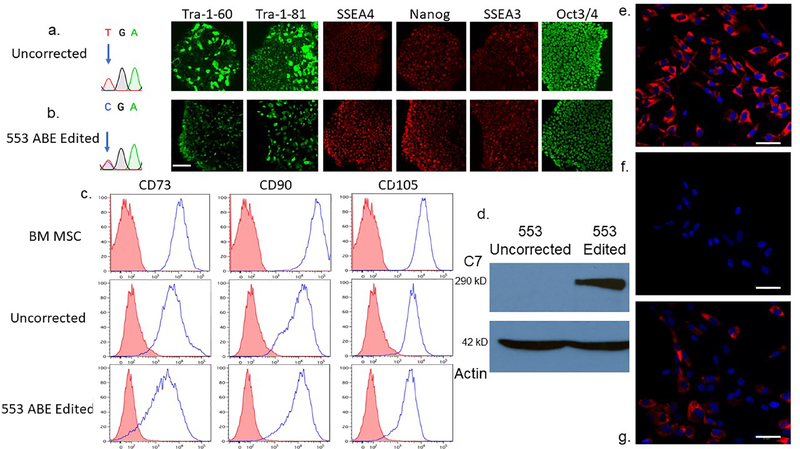
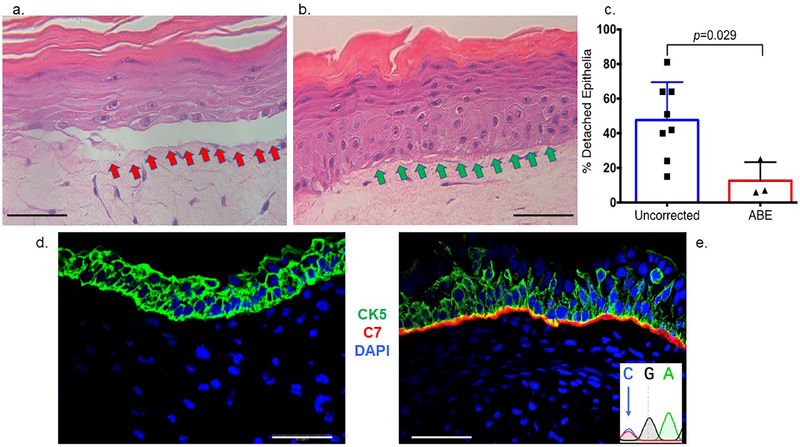
Genome editing represents a promising strategy for the therapeutic correction of COL7A1 mutations that cause recessive dystrophic epidermolysis bullosa (RDEB). DNA cleavage followed by homology-directed repair (HDR) using an exogenous template has previously been used to correct COL7A1 mutations. HDR rates can be modest, and the double-strand DNA breaks that initiate HDR commonly result in accompanying undesired insertions and deletions (indels). To overcome these limitations, we applied an A•T→G•C adenine base editor (ABE) to correct two different COL7A1 mutations in primary fibroblasts derived from RDEB patients. ABE enabled higher COL7A1 correction efficiencies than previously reported HDR efforts. Moreover, ABE obviated the need for a repair template, and minimal indels or editing at off-target sites was detected. Base editing restored the endogenous type VII collagen expression and function in vitro. We also treated induced pluripotent stem cells (iPSCs) derived from RDEB fibroblasts with ABE. The edited iPSCs were differentiated into mesenchymal stromal cells, a cell population with therapeutic potential for RDEB. In a mouse teratoma model, the skin derived from ABE-treated iPSCs showed the proper deposition of C7 at the dermal-epidermal junction in vivo. These demonstrate that base editing provides an efficient and precise genome editing method for autologous cell engineering for RDEB.
Copyright © 2019 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
CONFLICT OF INTEREST.
G.A.N. has filed patents relating to base editor use.
D.R.L. is a consultant and cofounder of Editas Medicine, Pairwise Plants, and Beam Therapeutics, companies that use genome editing.
B.R.B. is cofounder of Tmunity.
Figures
References
-
- Chen M, Costa FK, Lindvay CR, Han YP, Woodley DT. The recombinant expression of full-length type VII collagen and characterization of molecular mechanisms underlying dystrophic epidermolysis bullosa. J Biol Chem 2002;277(3):2118–24. - PubMed
-
- Christiano AM, Anton-Lamprecht I, Amano S, Ebschner U, Burgeson RE, Uitto J. Compound heterozygosity for COL7A1 mutations in twins with dystrophic epidermolysis bullosa: a recessive paternal deletion/insertion mutation and a dominant negative maternal glycine substitution result in a severe phenotype. Am J Hum Genet 1996;58(4):682–93. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
