

The vault RNA of Trypanosoma brucei plays a role in the production of trans-spliced mRNA
- PMID: 31439669
- PMCID: PMC6816085
- DOI: 10.1074/jbc.RA119.008580
The vault RNA of Trypanosoma brucei plays a role in the production of trans-spliced mRNA
Abstract
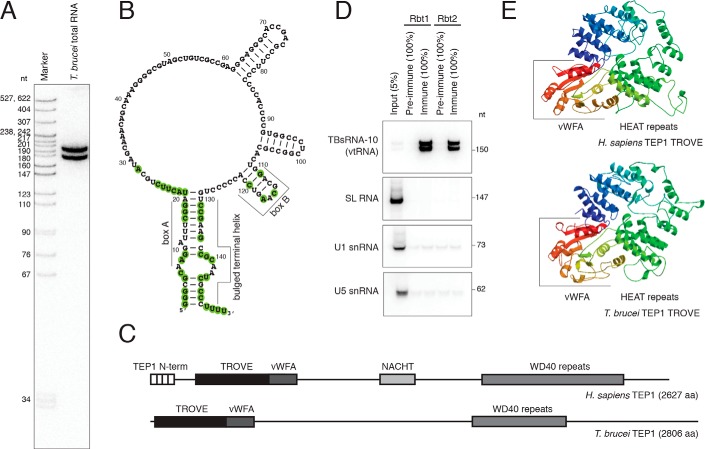
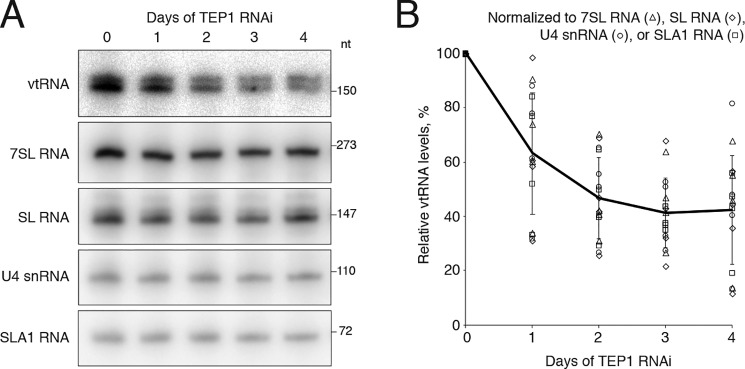
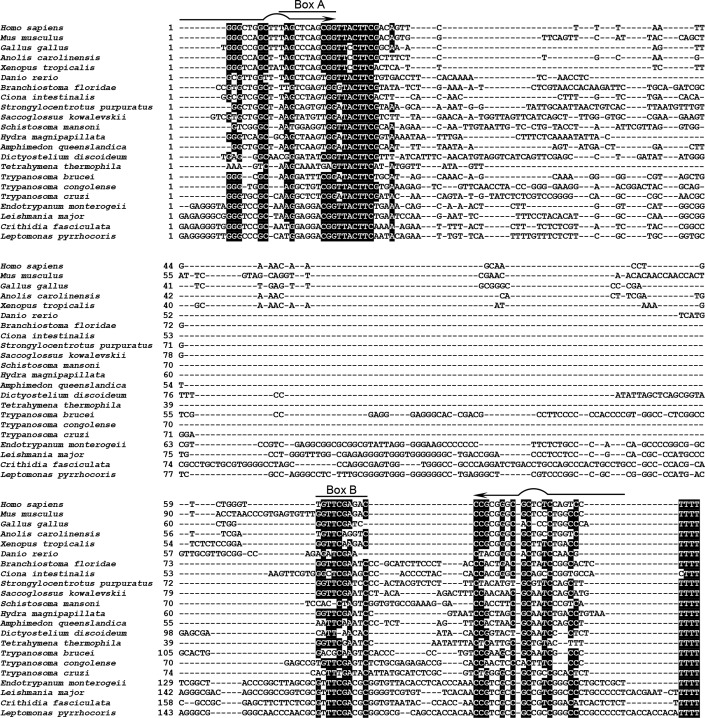
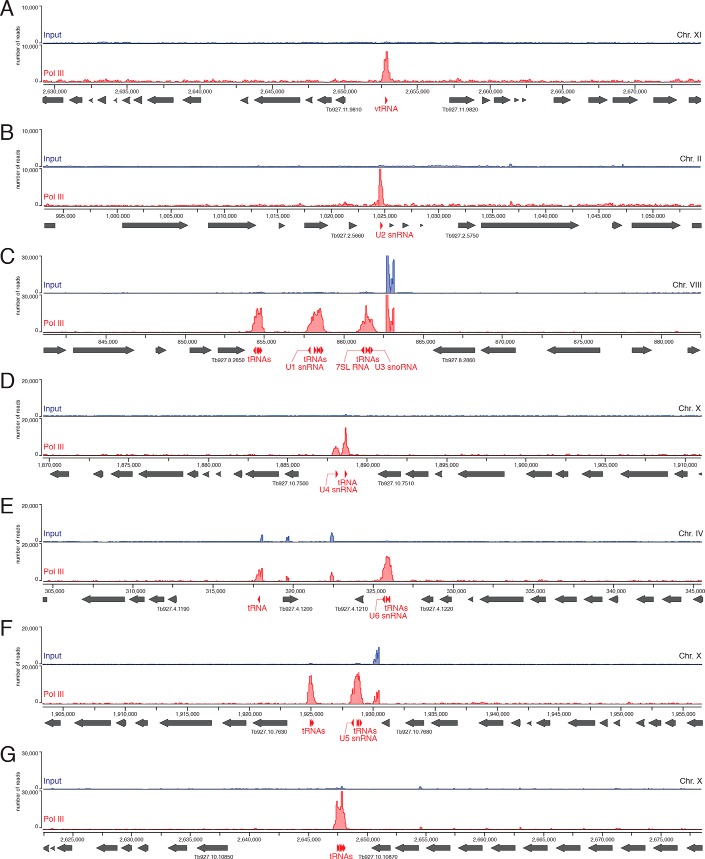
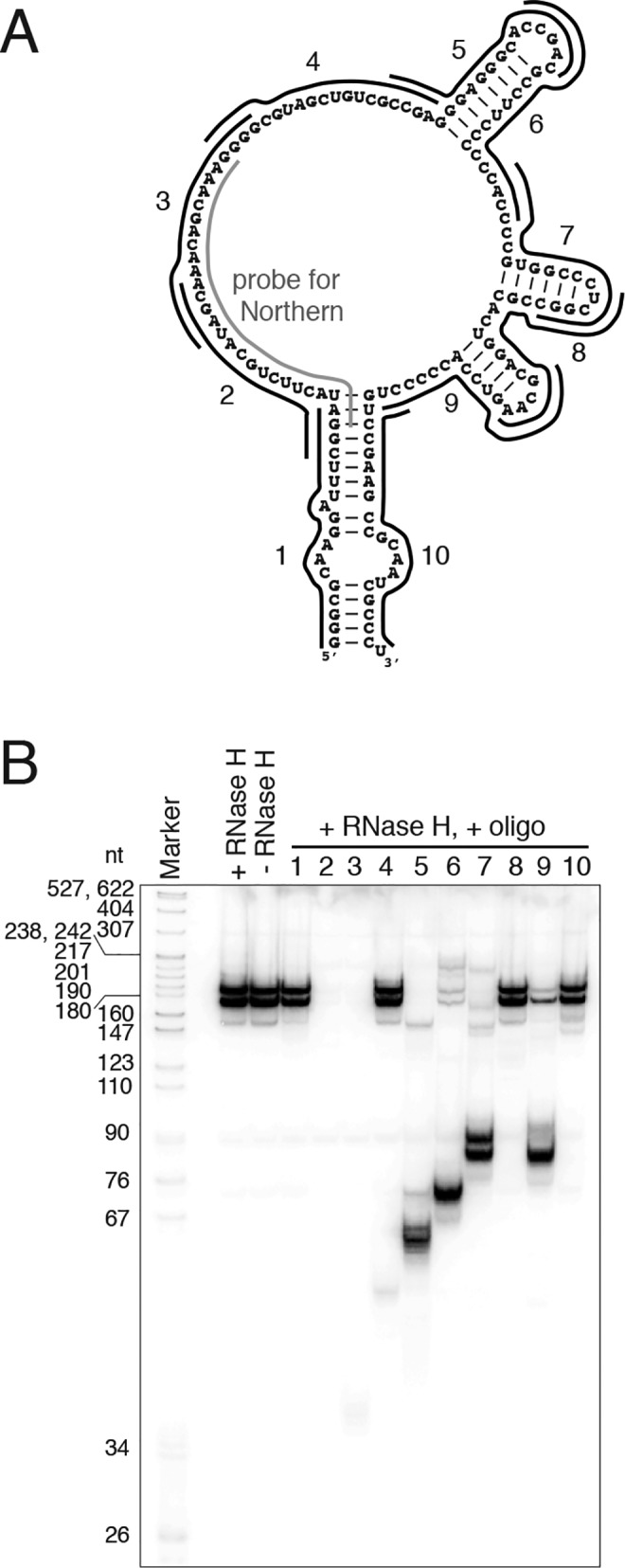
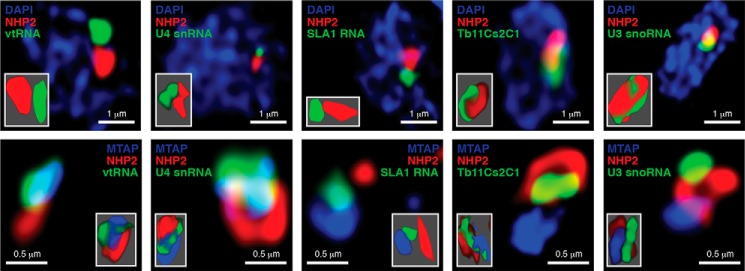
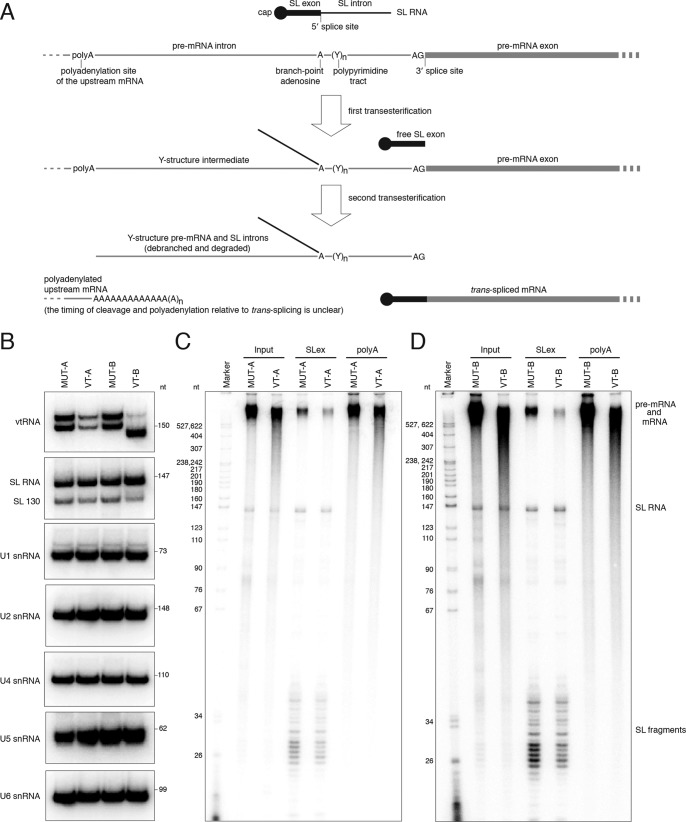
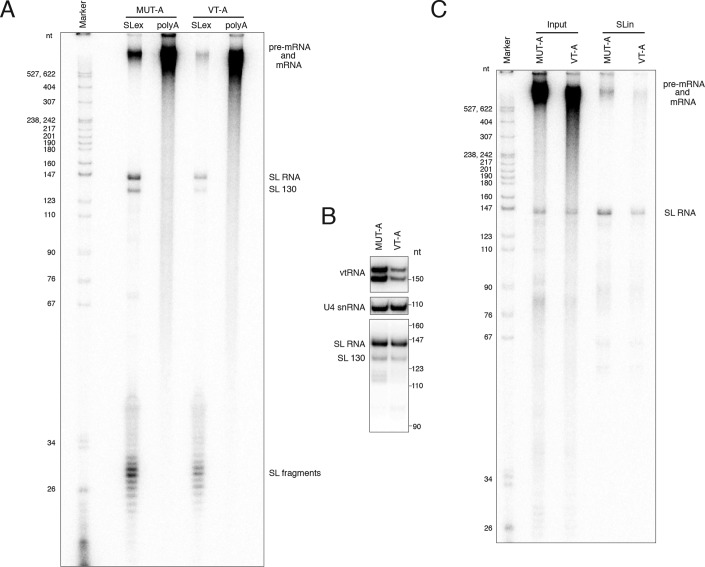
The vault ribonucleoprotein (RNP), comprising vault RNA (vtRNA) and telomerase-associated protein 1 (TEP1), is found in many eukaryotes. However, previous studies of vtRNAs, for example in mammalian cells, have failed to reach a definitive conclusion about their function. vtRNAs are related to Y RNAs, which are complexed with Ro protein and influence Ro's function in noncoding RNA (ncRNA) quality control and processing. In Trypanosoma brucei, the small noncoding TBsRNA-10 was first described in a survey of the ncRNA repertoire in this organism. Here, we report that TBsRNA-10 in T. brucei is a vtRNA, based on its association with TEP1 and sequence similarity to those of other known and predicted vtRNAs. We observed that like vtRNAs in other species, TBsRNA-10 is transcribed by RNA polymerase III, which in trypanosomes also generates the spliceosomal U-rich small nuclear RNAs. In T. brucei, spliced leader (SL)-mediated trans-splicing of pre-mRNAs is an obligatory step in gene expression, and we found here that T. brucei's vtRNA is highly enriched in a non-nucleolar locus in the cell nucleus implicated in SL RNP biogenesis. Using a newly developed permeabilized cell system for the bloodstream form of T. brucei, we show that down-regulated vtRNA levels impair trans-spliced mRNA production, consistent with a role of vtRNA in trypanosome mRNA metabolism. Our results suggest a common theme for the functions of vtRNAs and Y RNAs. We conclude that by complexing with their protein-binding partners TEP1 and Ro, respectively, these two RNA species modulate the metabolism of various RNA classes.
Keywords: RNA; RNA metabolism; RNA processing; RNA splicing; RNA-binding protein; Ro protein; Trypanosoma brucei; spliced leader (SL); telomerase-associated protein 1 (TEP1); vault RNA.
© 2019 Kolev et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
Comment in
-
A new role for vault RNA-TEP1 complexes in mRNA production in trypanosomes.J Biol Chem. 2019 Oct 25;294(43):15575-15576. doi: 10.1074/jbc.H119.011130. J Biol Chem. 2019. PMID: 31653772 Free PMC article.
References
-
- Kickhoefer V. A., Searles R. P., Kedersha N. L., Garber M. E., Johnson D. L., and Rome L. H. (1993) Vault ribonucleoprotein particles from rat and bullfrog contain a related small RNA that is transcribed by RNA polymerase III. J. Biol. Chem. 268, 7868–7873 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
