

Kinetic asymmetry allows macromolecular catalysts to drive an information ratchet
- PMID: 31444340
- PMCID: PMC6707331
- DOI: 10.1038/s41467-019-11402-7
Kinetic asymmetry allows macromolecular catalysts to drive an information ratchet
Abstract
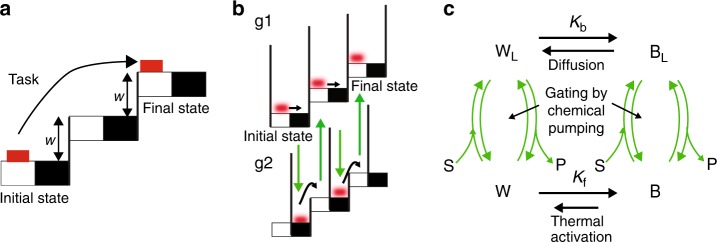
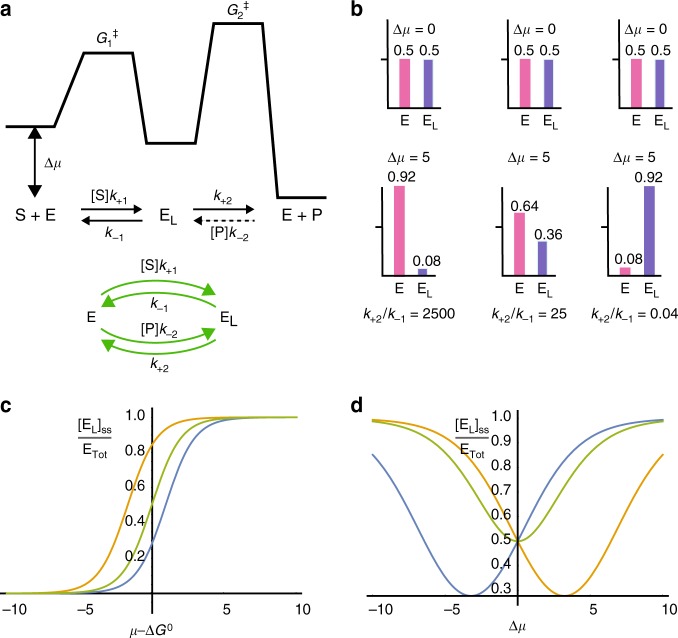
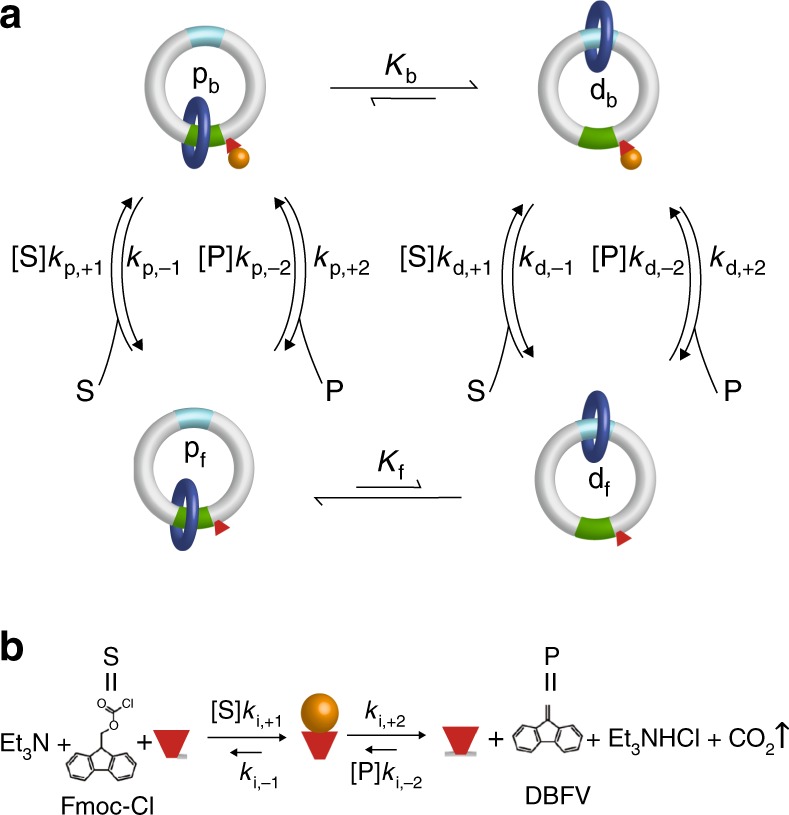
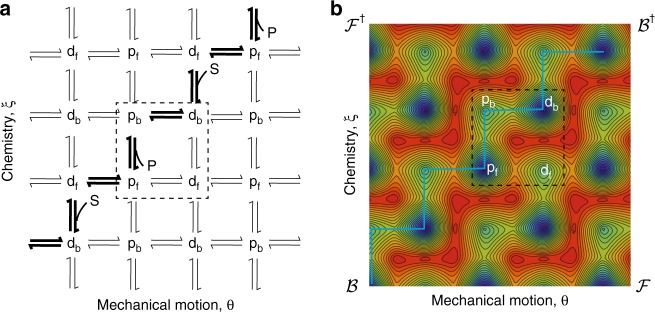
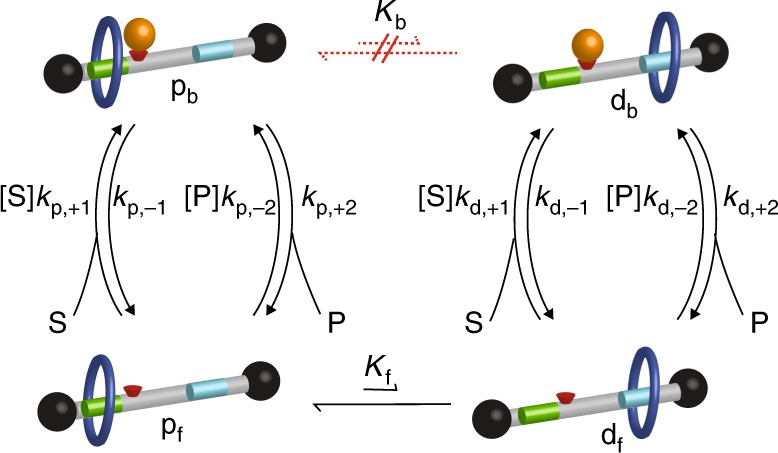
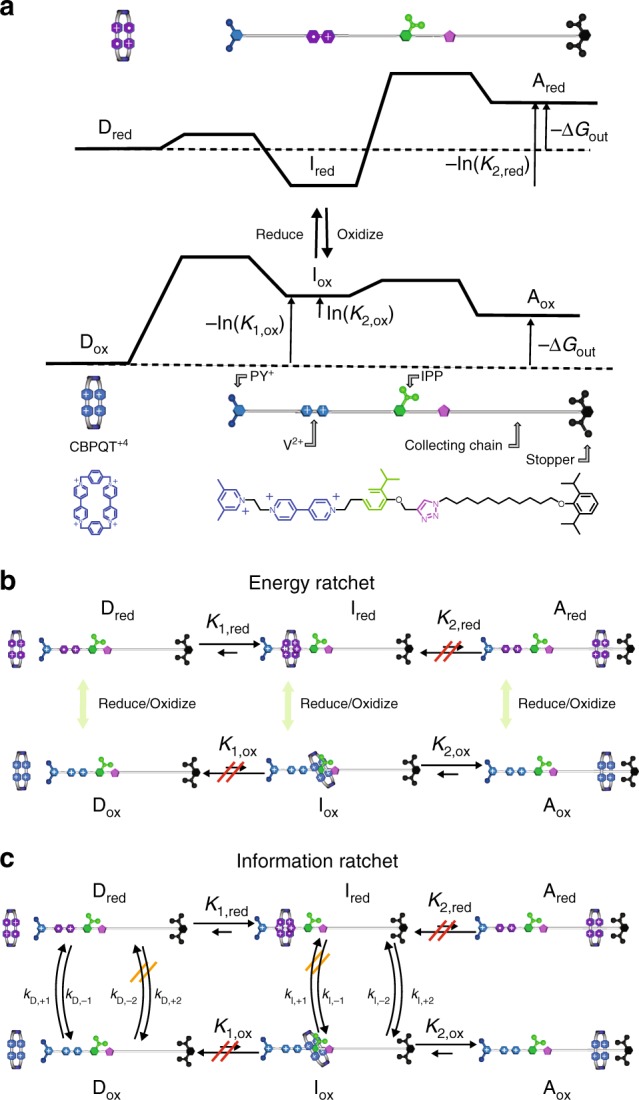
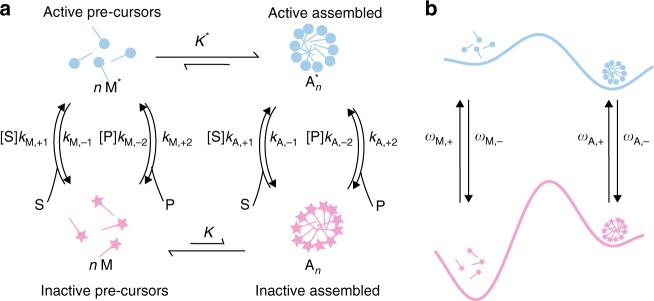
Molecular machines carry out their function by equilibrium mechanical motions in environments that are far from thermodynamic equilibrium. The mechanically equilibrated character of the trajectories of the macromolecule has allowed development of a powerful theoretical description, reminiscent of Onsager's trajectory thermodynamics, that is based on the principle of microscopic reversibility. Unlike the situation at thermodynamic equilibrium, kinetic parameters play a dominant role in determining steady-state concentrations away from thermodynamic equilibrium, and kinetic asymmetry provides a mechanism by which chemical free-energy released by catalysis can drive directed motion, molecular adaptation, and self-assembly. Several examples drawn from the recent literature, including a catenane-based chemically driven molecular rotor and a synthetic molecular assembler or pump, are discussed.
Conflict of interest statement
The author declares no competing interests.
Figures
References
-
- Ragazzon G, Prins LJ. Energy consumption in chemical fuel-driven self-assembly. Nat. Nanotechnol. 2018;11:1. - PubMed
Publication types
LinkOut - more resources
Full Text Sources
