

Aberrant Function of the C-Terminal Tail of HIST1H1E Accelerates Cellular Senescence and Causes Premature Aging
- PMID: 31447100
- PMCID: PMC6731364
- DOI: 10.1016/j.ajhg.2019.07.007
Aberrant Function of the C-Terminal Tail of HIST1H1E Accelerates Cellular Senescence and Causes Premature Aging
Abstract
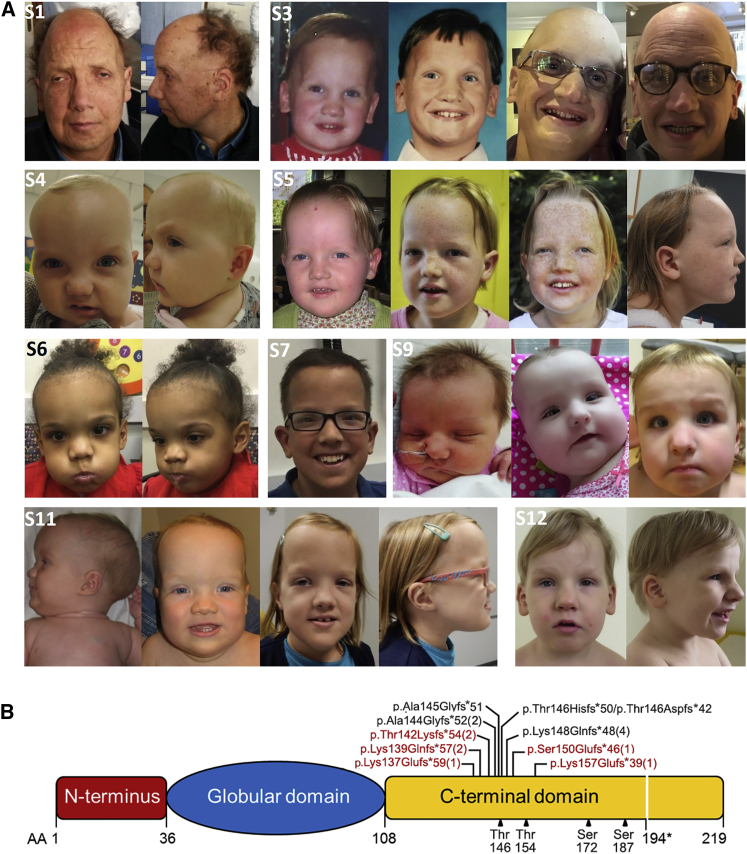
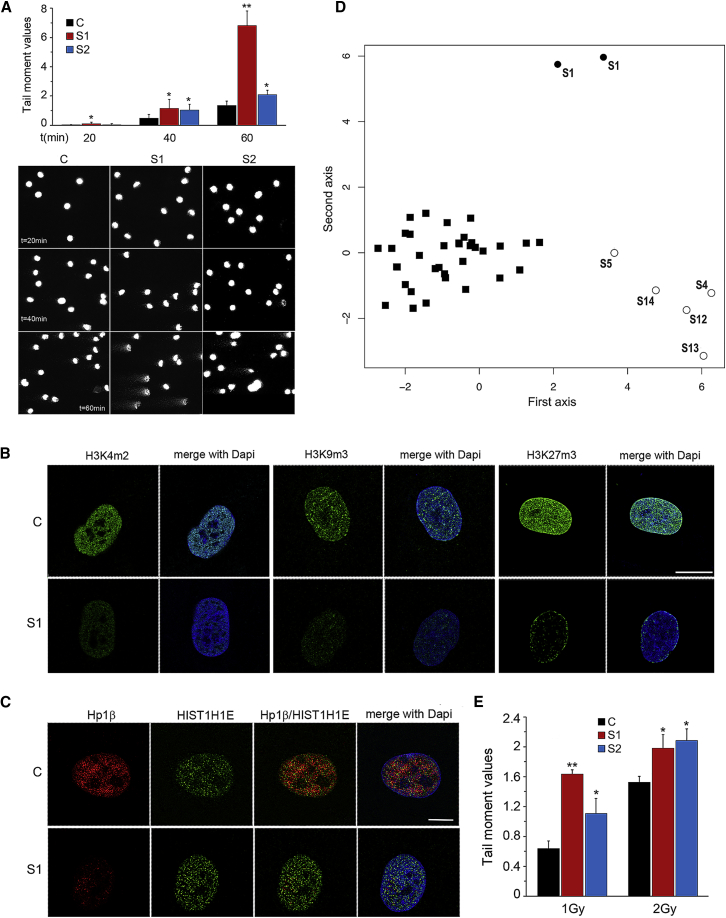
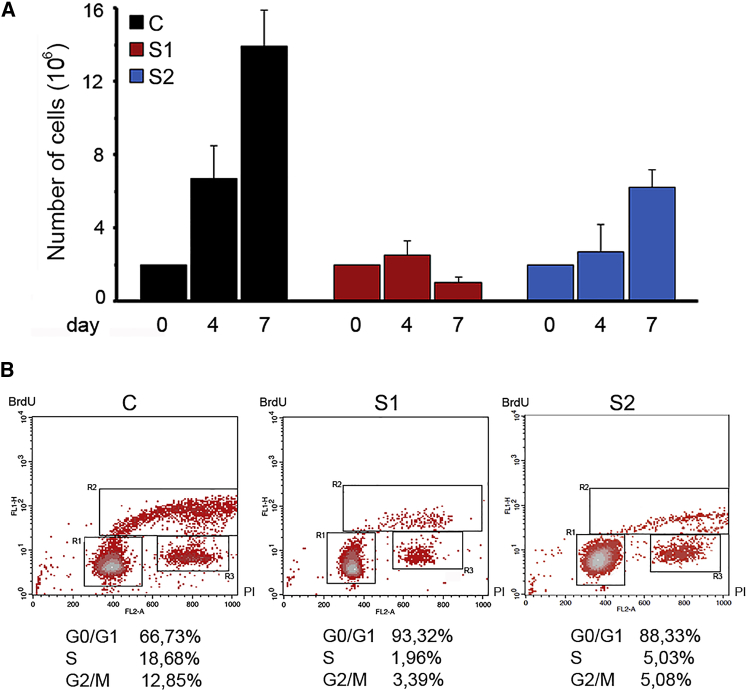
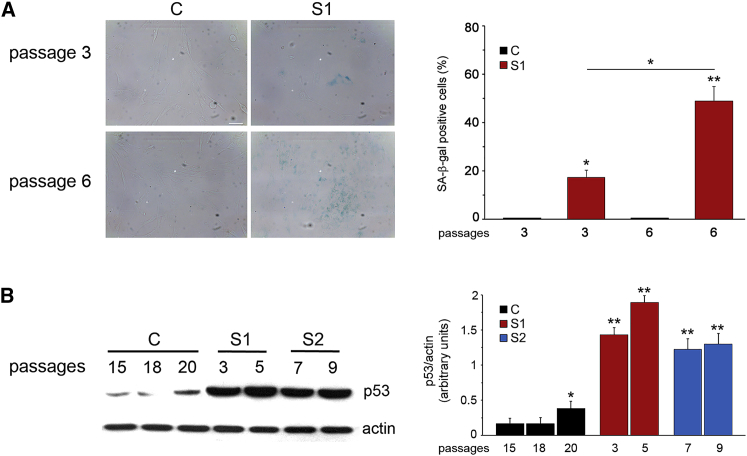
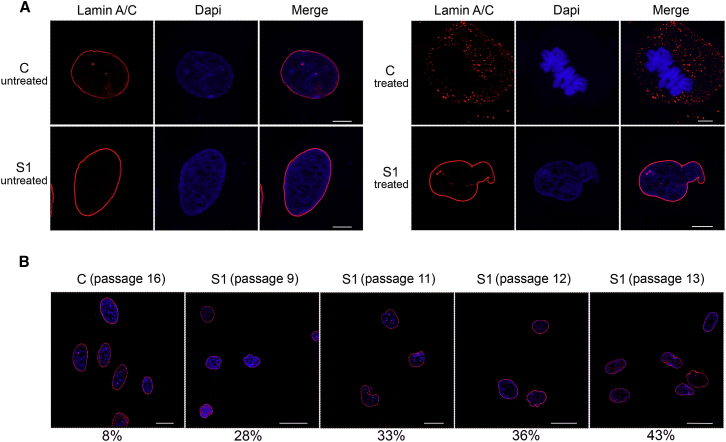
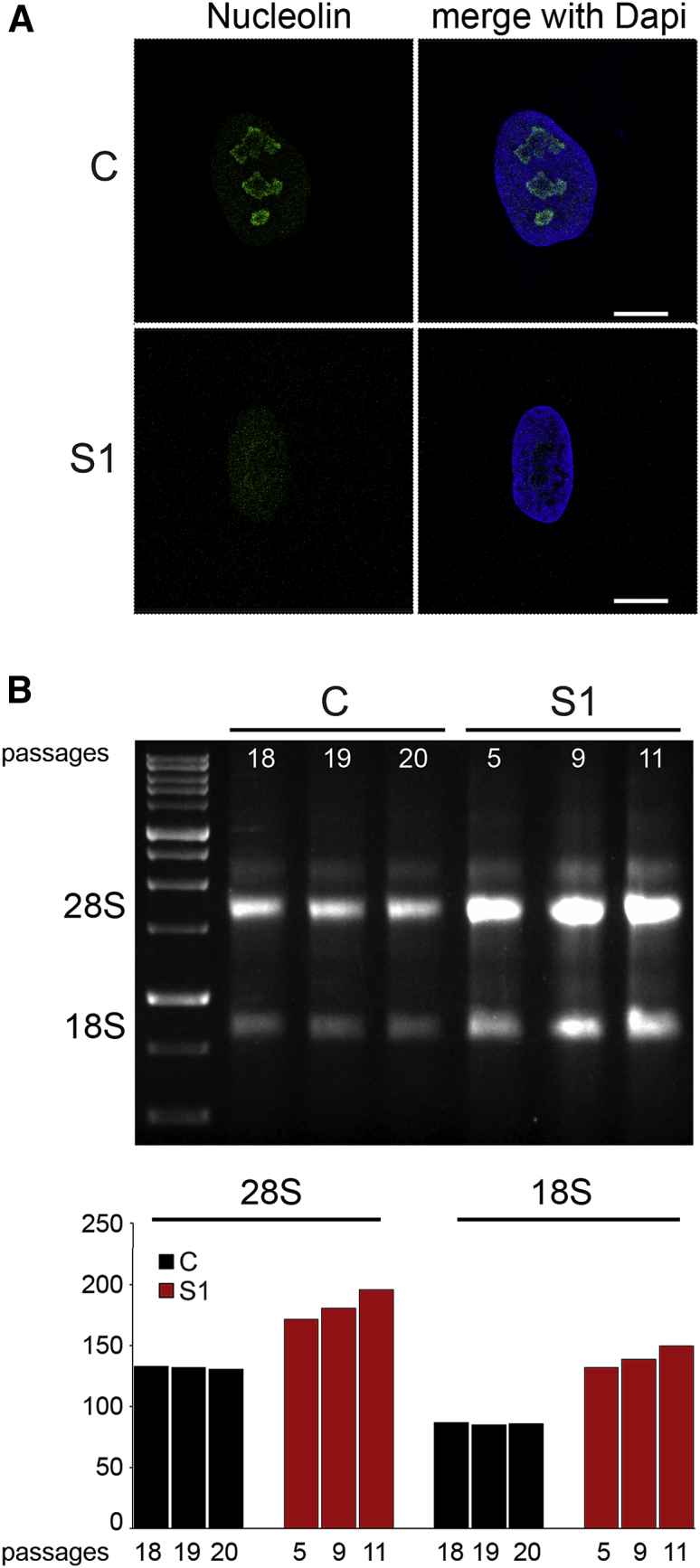
Histones mediate dynamic packaging of nuclear DNA in chromatin, a process that is precisely controlled to guarantee efficient compaction of the genome and proper chromosomal segregation during cell division and to accomplish DNA replication, transcription, and repair. Due to the important structural and regulatory roles played by histones, it is not surprising that histone functional dysregulation or aberrant levels of histones can have severe consequences for multiple cellular processes and ultimately might affect development or contribute to cell transformation. Recently, germline frameshift mutations involving the C-terminal tail of HIST1H1E, which is a widely expressed member of the linker histone family and facilitates higher-order chromatin folding, have been causally linked to an as-yet poorly defined syndrome that includes intellectual disability. We report that these mutations result in stable proteins that reside in the nucleus, bind to chromatin, disrupt proper compaction of DNA, and are associated with a specific methylation pattern. Cells expressing these mutant proteins have a dramatically reduced proliferation rate and competence, hardly enter into the S phase, and undergo accelerated senescence. Remarkably, clinical assessment of a relatively large cohort of subjects sharing these mutations revealed a premature aging phenotype as a previously unrecognized feature of the disorder. Our findings identify a direct link between aberrant chromatin remodeling, cellular senescence, and accelerated aging.
Keywords: HIST1H1E; accelerated aging; cellular senescence; chromatin compaction; chromatin dynamics; chromatin remodeling; linker histone; linker histone H1.4; methylation profiling; replicative senescence.
Copyright © 2019 American Society of Human Genetics. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
A.Be. and R.P. declare no additional conflicts of interest beyond their employment affiliation. All of the other authors declare no competing interests.
Figures
References
-
- Wolffe A. Academic Press; London: 1998. Chromatin: Structure and function.
-
- Luger K., Mäder A.W., Richmond R.K., Sargent D.F., Richmond T.J. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature. 1997;389:251–260. - PubMed
-
- Crane-Robinson C. Linker histones: History and current perspectives. Biochim. Biophys. Acta. 2016;1859:431–435. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
