

Epicardium in Heart Development
- PMID: 31451510
- PMCID: PMC6996450
- DOI: 10.1101/cshperspect.a037192
Epicardium in Heart Development
Abstract
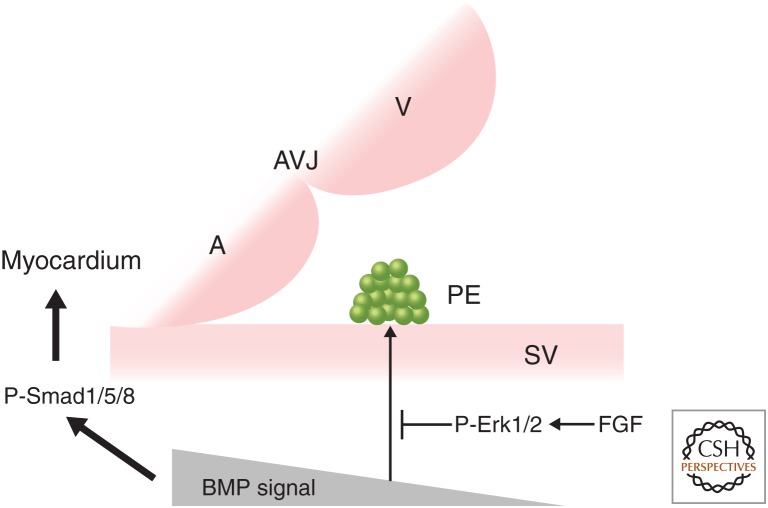
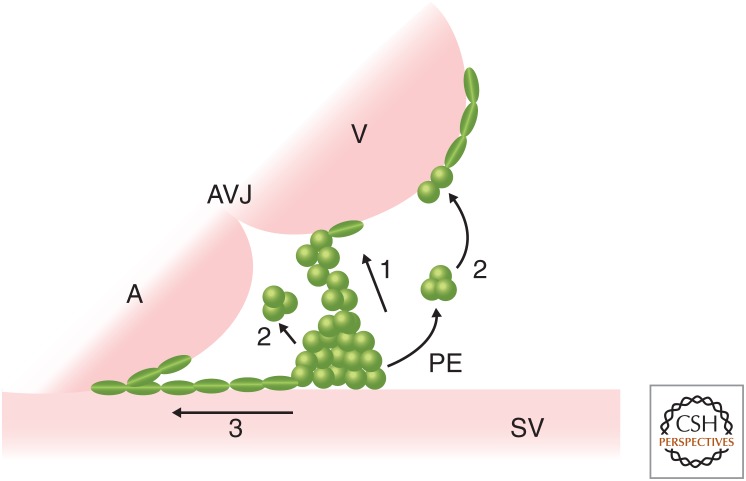
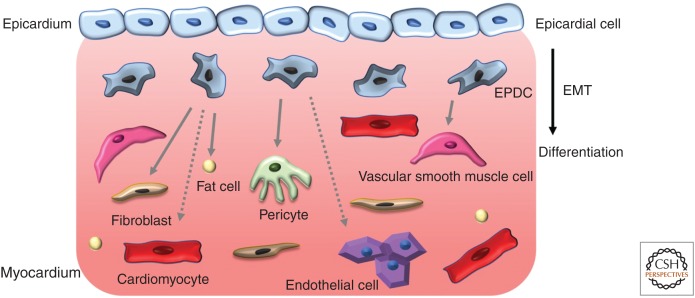
The epicardium, the outermost tissue layer that envelops all vertebrate hearts, plays a crucial role in cardiac development and regeneration and has been implicated in potential strategies for cardiac repair. The heterogenous cell population that composes the epicardium originates primarily from a transient embryonic cell cluster known as the proepicardial organ (PE). Characterized by its high cellular plasticity, the epicardium contributes to both heart development and regeneration in two critical ways: as a source of progenitor cells and as a critical signaling hub. Despite this knowledge, there are many unanswered questions in the field of epicardial biology, the resolution of which will advance the understanding of cardiac development and repair. We review current knowledge in cross-species epicardial involvement, specifically in relation to lineage specification and differentiation during cardiac development.
Copyright © 2020 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources