

Structural Biology of Telomerase
- PMID: 31451513
- PMCID: PMC6886448
- DOI: 10.1101/cshperspect.a032383
Structural Biology of Telomerase
Abstract
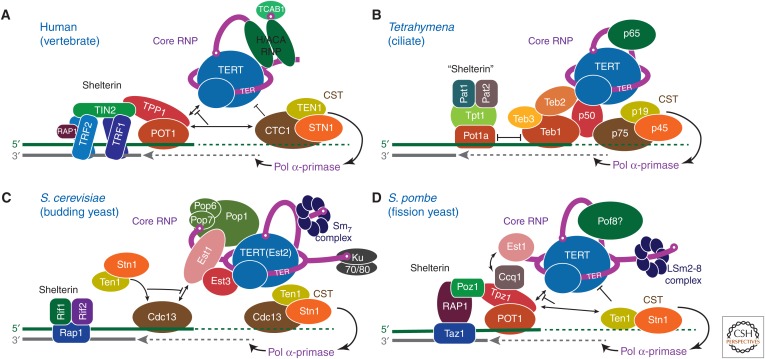
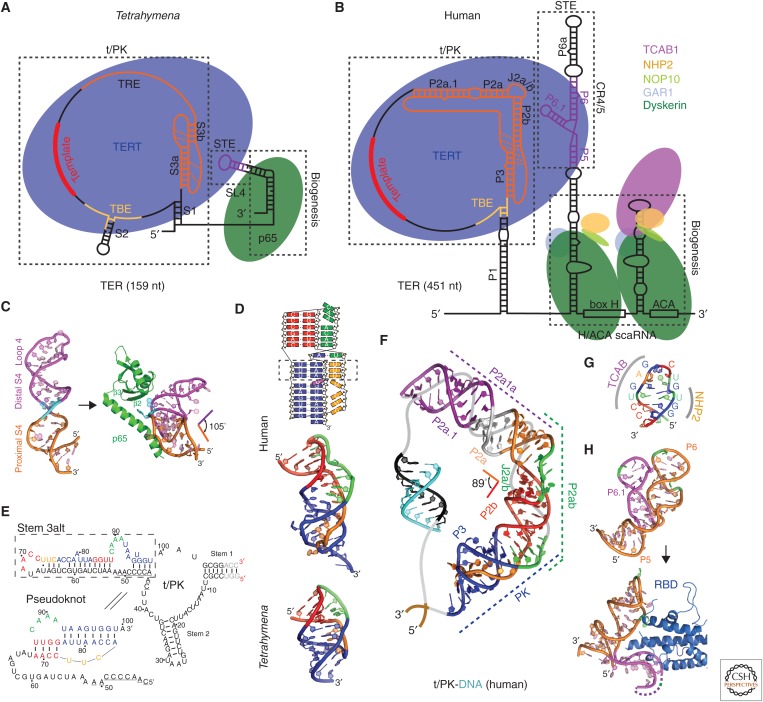
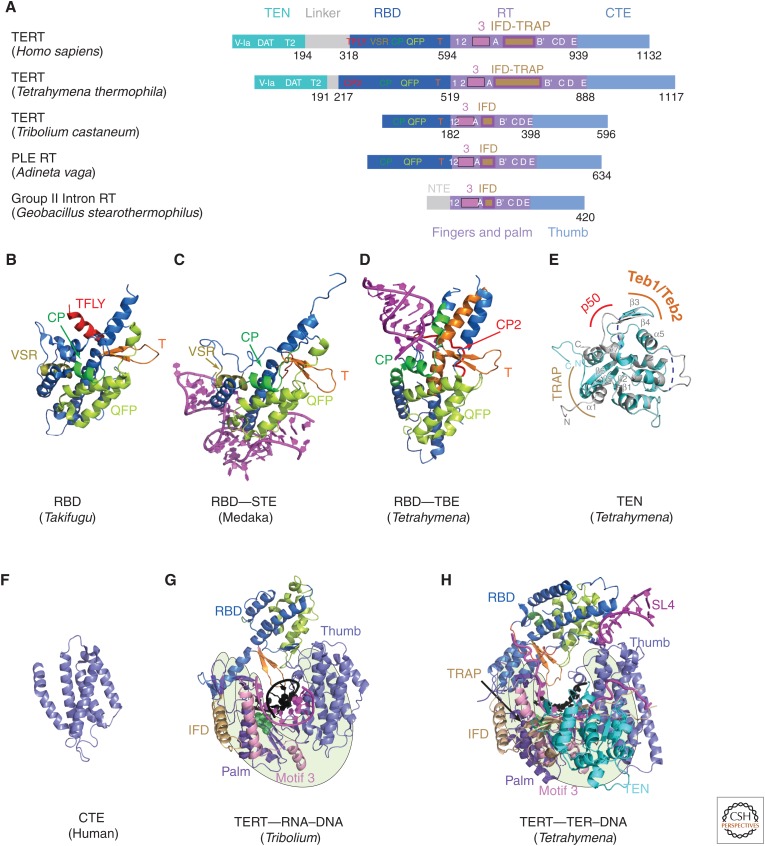
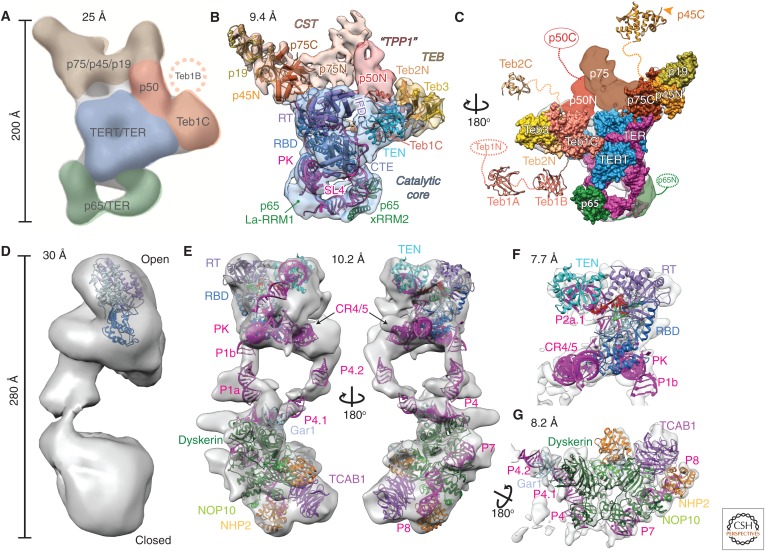
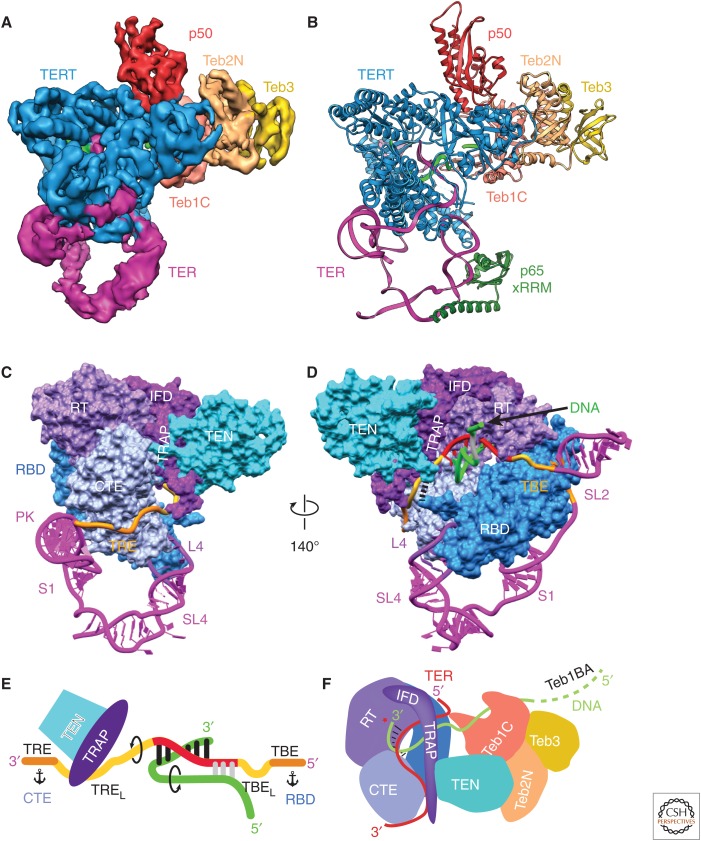
Telomerase is a DNA polymerase that extends the 3' ends of chromosomes by processively synthesizing multiple telomeric repeats. It is a unique ribonucleoprotein (RNP) containing a specialized telomerase reverse transcriptase (TERT) and telomerase RNA (TER) with its own template and other elements required with TERT for activity (catalytic core), as well as species-specific TER-binding proteins important for biogenesis and assembly (core RNP); other proteins bind telomerase transiently or constitutively to allow association of telomerase and other proteins with telomere ends for regulation of DNA synthesis. Here we describe how nuclear magnetic resonance (NMR) spectroscopy and X-ray crystallography of TER and protein domains helped define the structure and function of the core RNP, laying the groundwork for interpreting negative-stain and cryo electron microscopy (cryo-EM) density maps of Tetrahymena thermophila and human telomerase holoenzymes. As the resolution has improved from ∼30 Å to ∼5 Å, these studies have provided increasingly detailed information on telomerase architecture and mechanism.
Copyright © 2019 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
Similar articles
-
The architecture of Tetrahymena telomerase holoenzyme.Nature. 2013 Apr 11;496(7444):187-92. doi: 10.1038/nature12062. Epub 2013 Apr 3. Nature. 2013. PMID: 23552895 Free PMC article.
-
Structural Biology of Telomerase and Associated Factors.Cold Spring Harb Perspect Biol. 2025 Apr 15:a041697. doi: 10.1101/cshperspect.a041697. Online ahead of print. Cold Spring Harb Perspect Biol. 2025. PMID: 40234053
-
Structures of telomerase at several steps of telomere repeat synthesis.Nature. 2021 May;593(7859):454-459. doi: 10.1038/s41586-021-03529-9. Epub 2021 May 12. Nature. 2021. PMID: 33981033 Free PMC article.
-
Progress in structural studies of telomerase.Curr Opin Struct Biol. 2014 Feb;24:115-24. doi: 10.1016/j.sbi.2014.01.008. Epub 2014 Feb 4. Curr Opin Struct Biol. 2014. PMID: 24508601 Free PMC article. Review.
-
Structural biology of telomerase and its interaction at telomeres.Curr Opin Struct Biol. 2017 Dec;47:77-87. doi: 10.1016/j.sbi.2017.06.010. Epub 2017 Jul 18. Curr Opin Struct Biol. 2017. PMID: 28732250 Free PMC article. Review.
Cited by
-
[Molecular and cellular mechanisms of ageing: modern knowledge (literature review)].Probl Endokrinol (Mosk). 2023 Nov 11;69(5):45-54. doi: 10.14341/probl13278. Probl Endokrinol (Mosk). 2023. PMID: 37968951 Free PMC article. Review. Russian.
-
RIOK2 transcriptionally regulates TRiC and dyskerin complexes to prevent telomere shortening.Nat Commun. 2024 Aug 20;15(1):7138. doi: 10.1038/s41467-024-51336-3. Nat Commun. 2024. PMID: 39164231 Free PMC article.
-
Cooperative Analysis of Structural Dynamics in RNA-Protein Complexes by Single-Molecule Förster Resonance Energy Transfer Spectroscopy.Molecules. 2020 Apr 28;25(9):2057. doi: 10.3390/molecules25092057. Molecules. 2020. PMID: 32354083 Free PMC article. Review.
-
Structural biology of human telomerase: progress and prospects.Biochem Soc Trans. 2021 Nov 1;49(5):1927-1939. doi: 10.1042/BST20200042. Biochem Soc Trans. 2021. PMID: 34623385 Free PMC article. Review.
-
Does AlphaFold2 model proteins' intracellular conformations? An experimental test using cross-linking mass spectrometry of endogenous ciliary proteins.Commun Biol. 2023 Apr 15;6(1):421. doi: 10.1038/s42003-023-04773-7. Commun Biol. 2023. PMID: 37061613 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials