

Destabilization of the Tumor-Inducing Plasmid from an Octopine-Type Agrobacterium tumefaciens Lineage Drives a Large Deletion in the Co-resident At Megaplasmid
- PMID: 31451548
- PMCID: PMC6778807
- DOI: 10.1534/g3.119.400554
Destabilization of the Tumor-Inducing Plasmid from an Octopine-Type Agrobacterium tumefaciens Lineage Drives a Large Deletion in the Co-resident At Megaplasmid
Abstract
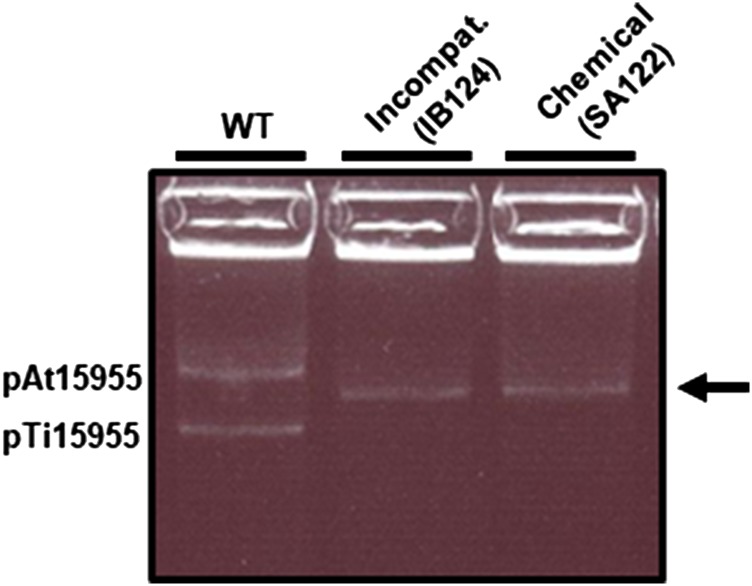
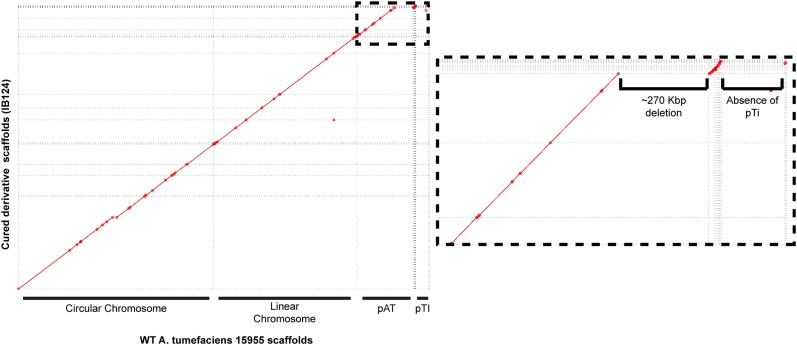
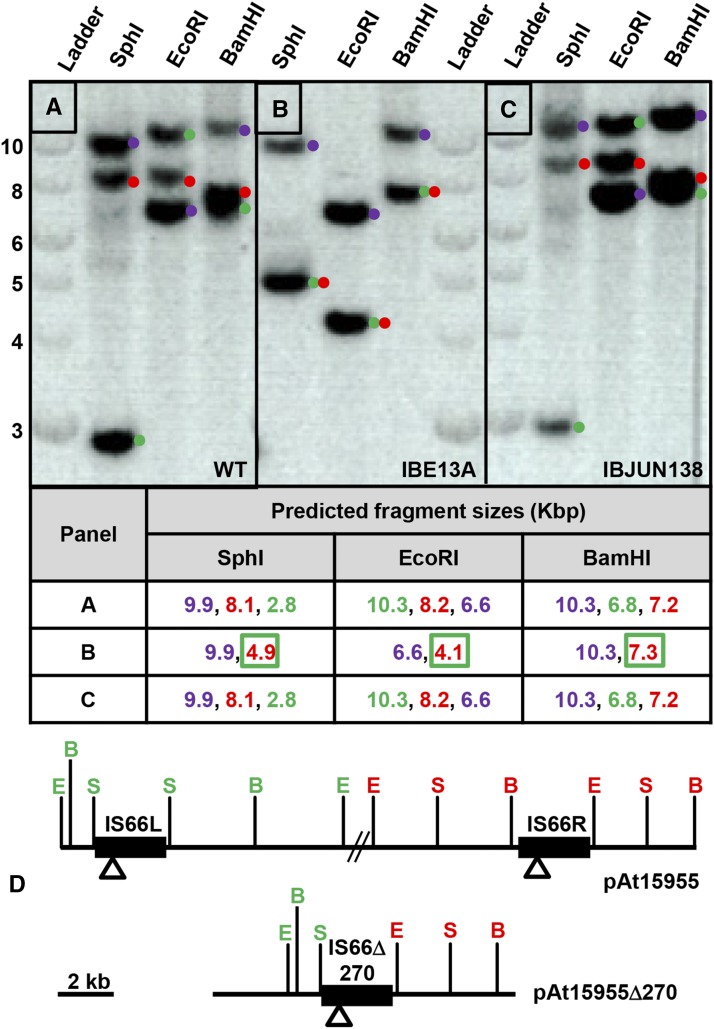
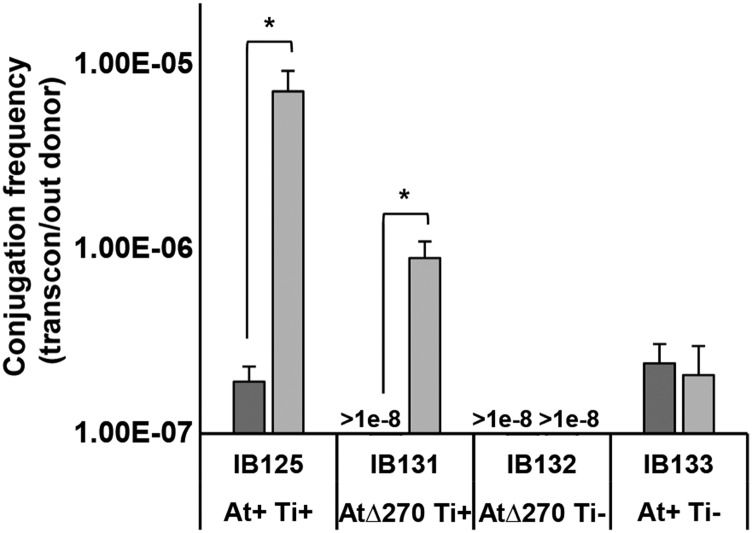
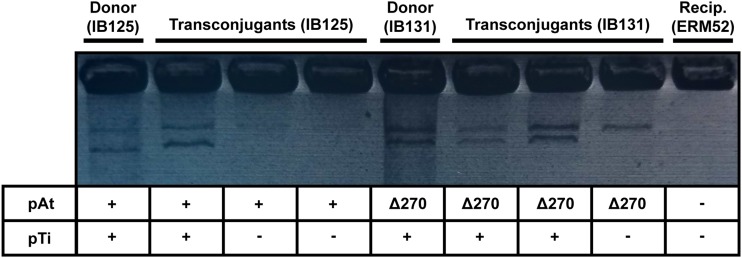
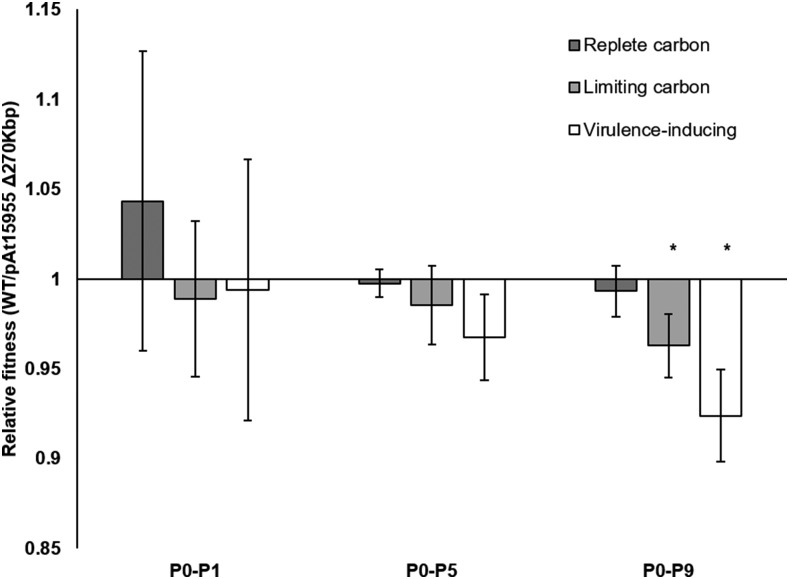
Bacteria with multi-replicon genome organizations, including members of the family Rhizobiaceae, often carry a variety of niche-associated functions on large plasmids. While evidence exists for cross-replicon interactions and co-evolution between replicons in many of these systems, remarkable strain-to-strain variation is also observed for extrachromosomal elements, suggesting increased genetic plasticity. Here, we show that curing of the tumor-inducing virulence plasmid (pTi) of an octopine-type Agrobacterium tumefaciens lineage leads to a large deletion in the co-resident At megaplasmid (pAt). The deletion event is mediated by a repetitive IS-element, IS66, and results in a variety of environment-dependent fitness consequences, including loss of independent conjugal transfer of the plasmid. Interestingly, a related and otherwise wild-type A. tumefaciens strain is missing exactly the same large pAt segment as the pAt deletion derivatives, suggesting a similar event over its natural history. Overall, the findings presented here uncover a novel genetic interaction between the two large plasmids of A. tumefaciens and provide evidence for cross-replicon integration and co-evolution of these plasmids.
Keywords: IS element; Ti plasmid; accessory genome; conjugal transfer; deletion; plasmid curing.
Copyright © 2019 Barton et al.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources