

CRISPR-Cas9 screens identify regulators of antibody-drug conjugate toxicity
- PMID: 31451760
- PMCID: PMC6911768
- DOI: 10.1038/s41589-019-0342-2
CRISPR-Cas9 screens identify regulators of antibody-drug conjugate toxicity
Abstract
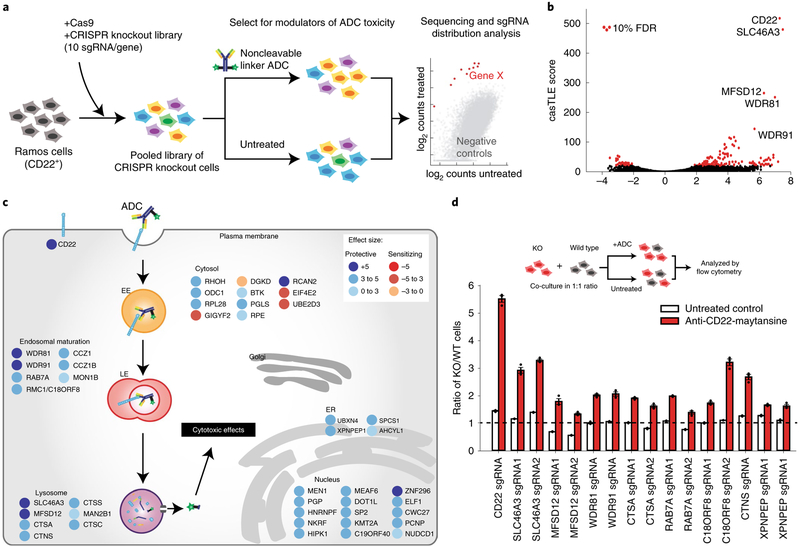
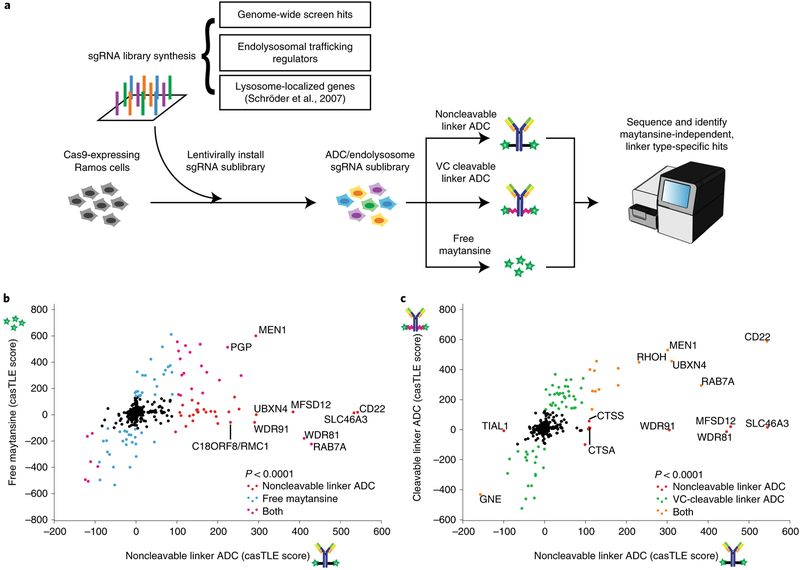
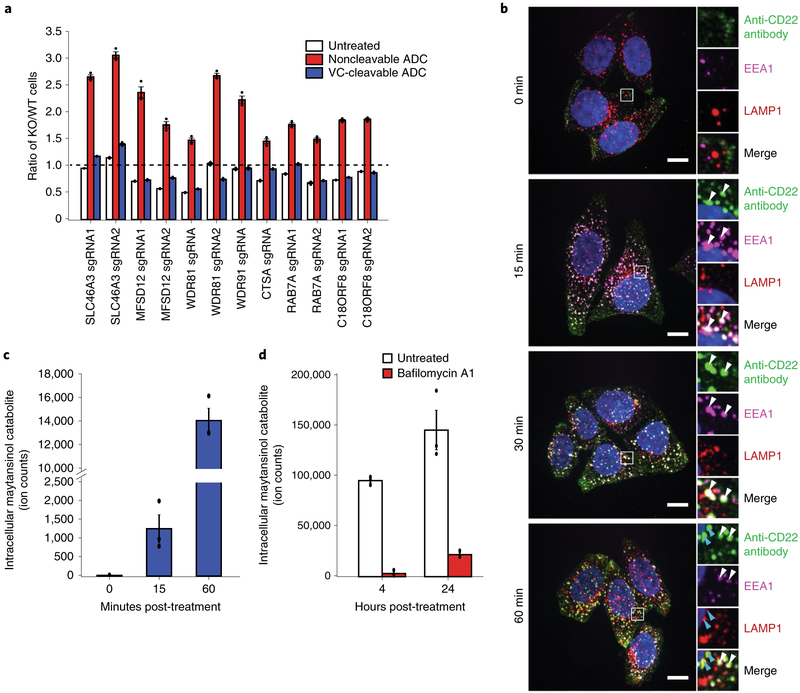
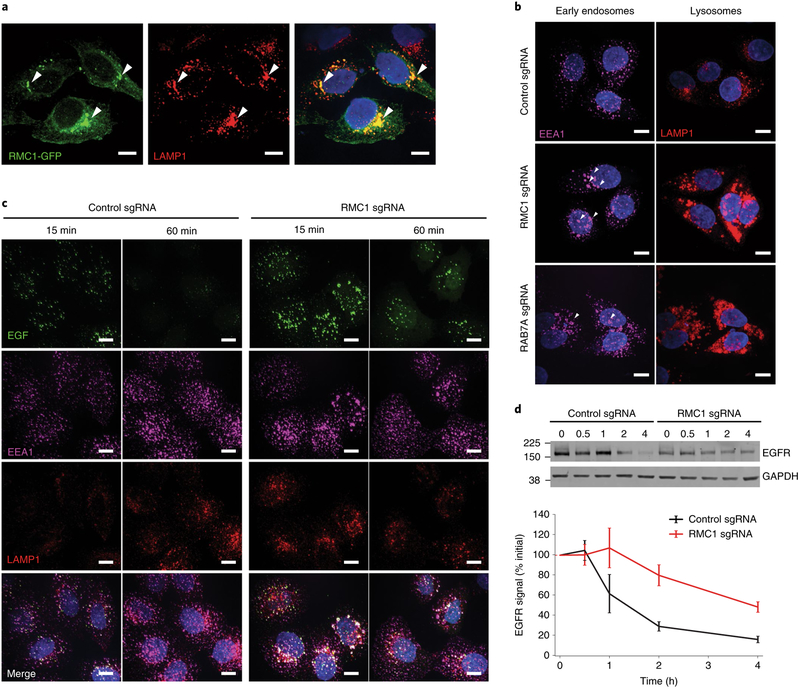
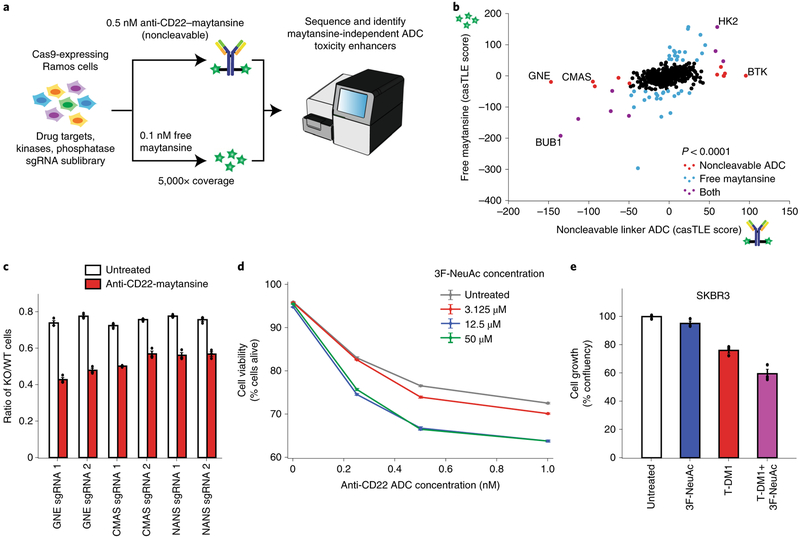
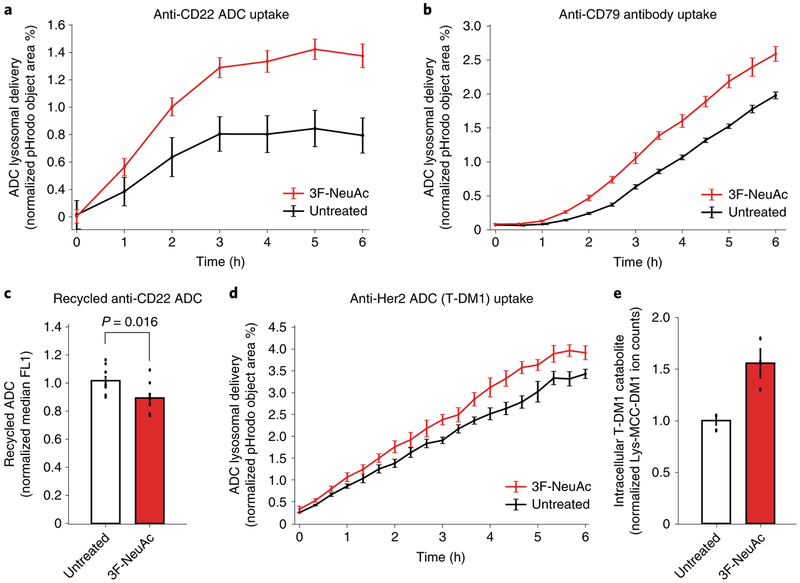
Antibody-drug conjugates (ADCs) selectively deliver chemotherapeutic agents to target cells and are important cancer therapeutics. However, the mechanisms by which ADCs are internalized and activated remain unclear. Using CRISPR-Cas9 screens, we uncover many known and novel endolysosomal regulators as modulators of ADC toxicity. We identify and characterize C18ORF8/RMC1 as a regulator of ADC toxicity through its role in endosomal maturation. Through comparative analysis of screens with ADCs bearing different linkers, we show that a subset of late endolysosomal regulators selectively influence toxicity of noncleavable linker ADCs. Surprisingly, we find cleavable valine-citrulline linkers can be processed rapidly after internalization without lysosomal delivery. Lastly, we show that sialic acid depletion enhances ADC lysosomal delivery and killing in diverse cancer cell types, including with FDA (US Food and Drug Administration)-approved trastuzumab emtansine (T-DM1) in Her2-positive breast cancer cells. Together, these results reveal new regulators of endolysosomal trafficking, provide important insights for ADC design and identify candidate combination therapy targets.
Conflict of interest statement
Competing interests
R.M.B. and D.R. are employees of Catalent Biologics. C.R.B. is a cofounder and Scientific Advisory Board member of Redwood Bioscience, which generated antibody-drug conjugates used in this work. Stanford University has filed a patent application based on the findings in this article.
Figures
References
-
- Beck A, Goetsch L, Dumontet C & Corvaïa N Strategies and challenges for the next generation of antibody-drug conjugates. Nat. Rev. Drug Discov 16, 315–337 (2017). - PubMed
-
- Lyon R Drawing lessons from the clinical development of antibody-drug conjugates. Drug Discov. Today Technol 30, 105–109 (2018). - PubMed
-
- Loganzo F et al. Tumor cells chronically treated with a trastuzumabmaytansinoid antibody-drug conjugate develop varied resistance mechanisms but respond to alternate treatments. Mol. Cancer Ther 14, 952–963 (2015). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
