

Isoproterenol enhances force production in mouse glycolytic and oxidative muscle via separate mechanisms
- PMID: 31451903
- PMCID: PMC6814637
- DOI: 10.1007/s00424-019-02304-0
Isoproterenol enhances force production in mouse glycolytic and oxidative muscle via separate mechanisms
Abstract
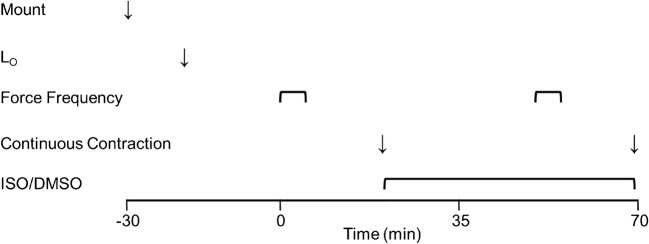
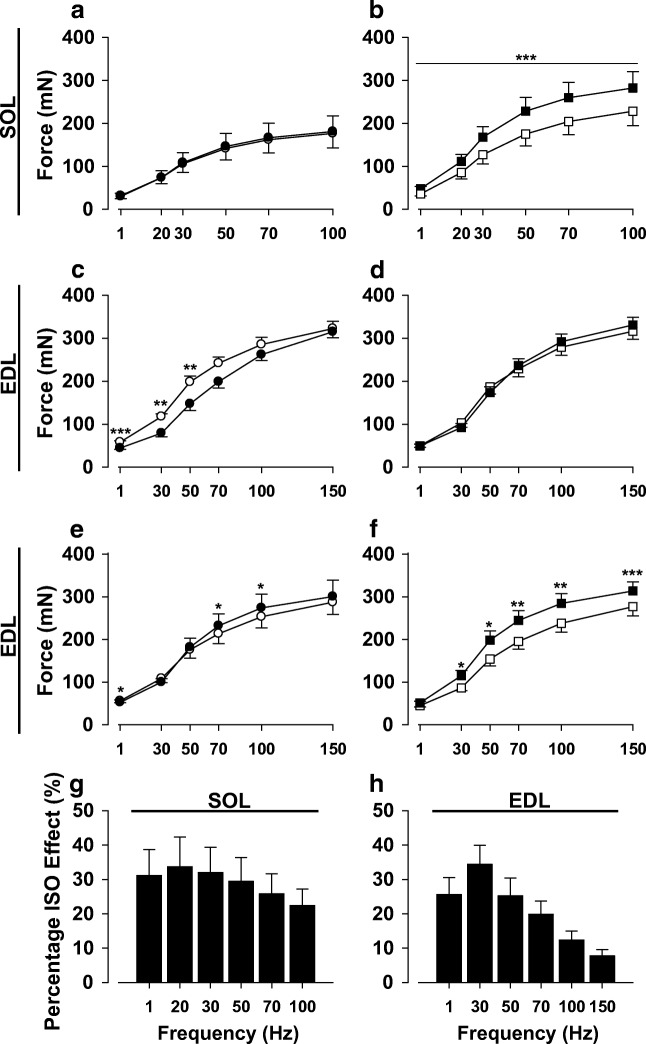
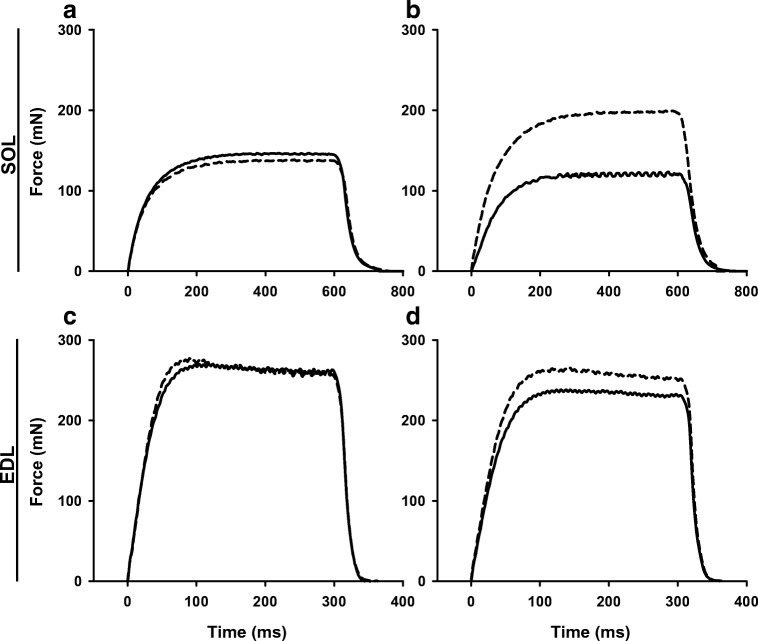
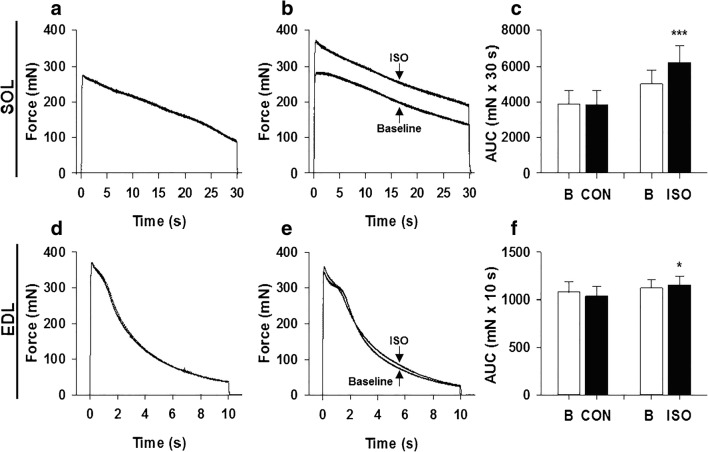
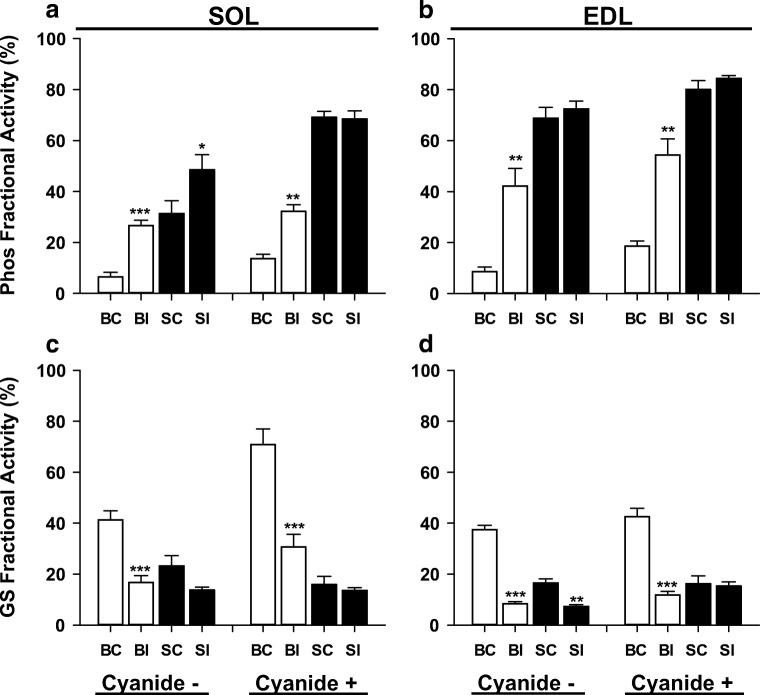
Fight or flight is a biologic phenomenon that involves activation of β-adrenoceptors in skeletal muscle. However, how force generation is enhanced through adrenergic activation in different muscle types is not fully understood. We studied the effects of isoproterenol (ISO, β-receptor agonist) on force generation and energy metabolism in isolated mouse soleus (SOL, oxidative) and extensor digitorum longus (EDL, glycolytic) muscles. Muscles were stimulated with isometric tetanic contractions and analyzed for metabolites and phosphorylase activity. Under conditions of maximal force production, ISO enhanced force generation markedly more in SOL (22%) than in EDL (8%). Similarly, during a prolonged tetanic contraction (30 s for SOL and 10 s for EDL), ISO-enhanced the force × time integral more in SOL (25%) than in EDL (3%). ISO induced marked activation of phosphorylase in both muscles in the basal state, which was associated with glycogenolysis (less in SOL than in EDL), and in EDL only, a significant decrease (16%) in inorganic phosphate (Pi). ATP turnover during sustained contractions (1 s EDL, 5 s SOL) was not affected by ISO in EDL, but essentially doubled in SOL. Under conditions of maximal stimulation, ISO has a minor effect on force generation in EDL that is associated with a decrease in Pi, whereas ISO has a marked effect on force generation in SOL that is associated with an increase in ATP turnover. Thus, phosphorylase functions as a phosphate trap in ISO-mediated force enhancement in EDL and as a catalyzer of ATP supply in SOL.
Keywords: Fatigue; Force; Isoproterenol; Metabolites; Muscle; Phosphorylase.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Variable cytoplasmic actin expression impacts the sensitivity of different dystrophin-deficient mdx skeletal muscles to eccentric contraction.FEBS J. 2019 Jul;286(13):2562-2576. doi: 10.1111/febs.14831. Epub 2019 Apr 11. FEBS J. 2019. PMID: 30942954 Free PMC article.
-
Phosphate increase during fatigue affects crossbridge kinetics in intact mouse muscle at physiological temperature.J Physiol. 2017 Jul 1;595(13):4317-4328. doi: 10.1113/JP273672. Epub 2017 May 8. J Physiol. 2017. PMID: 28332714 Free PMC article.
-
Lpaatδ/Agpat4 deficiency impairs maximal force contractility in soleus and alters fibre type in extensor digitorum longus muscle.Biochim Biophys Acta Mol Cell Biol Lipids. 2018 Jul;1863(7):700-711. doi: 10.1016/j.bbalip.2018.04.001. Epub 2018 Apr 6. Biochim Biophys Acta Mol Cell Biol Lipids. 2018. PMID: 29627383
-
Effects of isoproterenol on rubidium transport in slow- and fast-twitch muscles from euthyroid and hyperthyroid rats.Pflugers Arch. 1986 Oct;407(4):440-4. doi: 10.1007/BF00652631. Pflugers Arch. 1986. PMID: 3022229
-
Inotropic effects on mammalian skeletal muscle change with contraction frequency.Can J Physiol Pharmacol. 2003 Aug;81(8):753-8. doi: 10.1139/y03-031. Can J Physiol Pharmacol. 2003. PMID: 12897803 Review.
Cited by
-
Cardiovascular response to anticipatory and reactionary postural perturbations in young adults.Exp Physiol. 2023 Sep;108(9):1144-1153. doi: 10.1113/EP091173. Epub 2023 Jul 17. Exp Physiol. 2023. PMID: 37458232 Free PMC article.
-
The conventional isoproterenol-induced heart failure model does not consistently mimic the diaphragmatic dysfunction observed in patients.PLoS One. 2020 Jul 30;15(7):e0236923. doi: 10.1371/journal.pone.0236923. eCollection 2020. PLoS One. 2020. PMID: 32730329 Free PMC article.
-
The role of glycogen phosphorylase in glycogen biogenesis in skeletal muscle after exercise.Sports Med Health Sci. 2022 Nov 12;5(1):29-33. doi: 10.1016/j.smhs.2022.11.001. eCollection 2023 Mar. Sports Med Health Sci. 2022. PMID: 36994178 Free PMC article. Review.
-
MCU-complex-mediated mitochondrial calcium signaling is impaired in Barth syndrome.Hum Mol Genet. 2022 Feb 3;31(3):376-385. doi: 10.1093/hmg/ddab254. Hum Mol Genet. 2022. PMID: 34494107 Free PMC article.
-
A century of exercise physiology: key concepts in regulation of glycogen metabolism in skeletal muscle.Eur J Appl Physiol. 2022 Aug;122(8):1751-1772. doi: 10.1007/s00421-022-04935-1. Epub 2022 Mar 30. Eur J Appl Physiol. 2022. PMID: 35355125 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
