

IL-6-HaloTag® enables live-cell plasma membrane staining, flow cytometry, functional expression, and de-orphaning of recombinant odorant receptors
- PMID: 31453235
- PMCID: PMC6706138
- DOI: 10.14440/jbm.2017.206
IL-6-HaloTag® enables live-cell plasma membrane staining, flow cytometry, functional expression, and de-orphaning of recombinant odorant receptors
Abstract
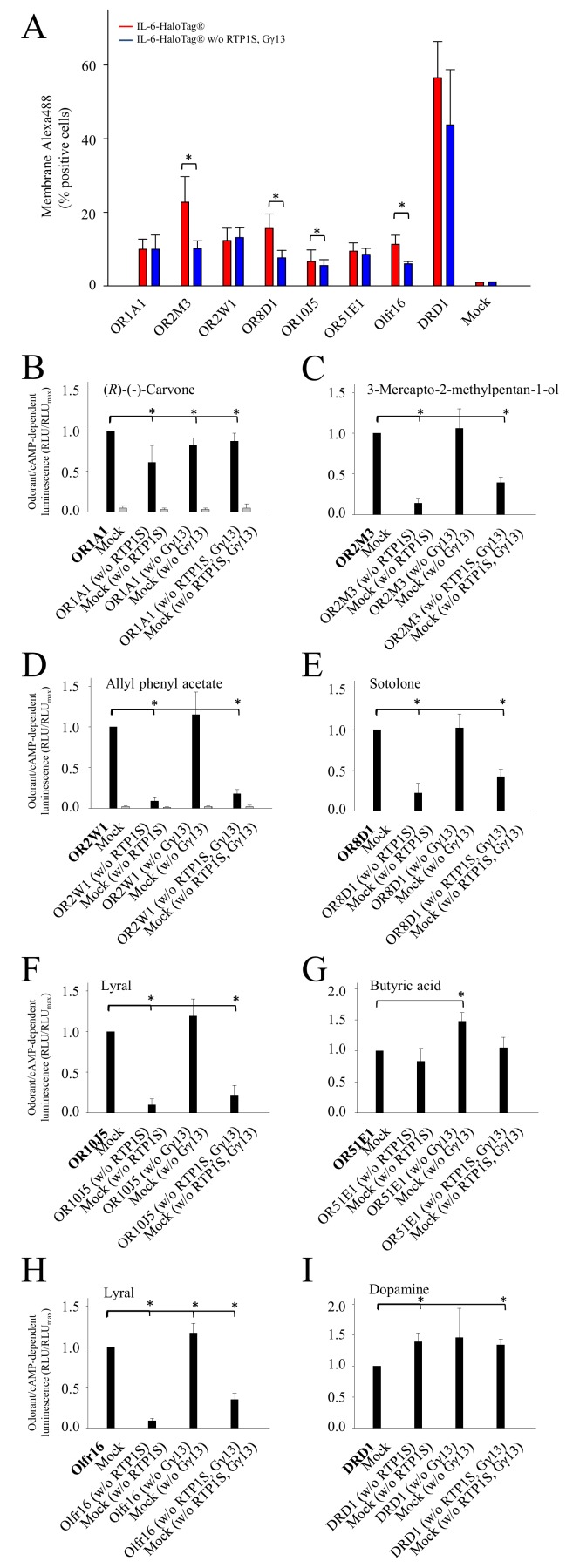
The assignment of cognate odorant/agonist pairs is a prerequisite for an understanding of odorant coding at the receptor level. However, the identification of new ligands for odorant receptors (ORs) in cell-based assays has been challenging, due to their individual and rather sub-optimal plasma membrane expression, as compared with other G protein-coupled receptors. Accessory proteins, such as the chaperone RTP1S, or Ric8b, have improved the surface expression of at least a portion of ORs. Typically, recombinant ORs carry N-terminal tags, which proved helpful for their functional membrane expression. The most common tag is the 'Rho-tag', representing an N-terminal part of rhodopsin, but also 'Lucy-' or 'Flag-tag' extensions have been described. Here, we used a bi-functional N-terminal tag, called 'interleukin 6 (IL-6)-HaloTag®', with IL-6 facilitating functional cell surface expression of recombinant ORs, and the HaloTag® protein, serving as a highly specific acceptor for cell-impermeant or cell-permeant, fluorophore-coupled ligands, which enable the quantification of odorant receptor expression by live-cell flow cytometry. Our experiments revealed on average an about four-fold increased surface expression, a four-fold higher signaling amplitude, and a significantly higher potency of odorant-induced cAMP signaling of six different human IL-6-HaloTag®-ORs across five different receptor families in NxG 108CC15 cells, as compared to their Rho-tag-HaloTag® constructs. We observed similar results in HEK-293 cells. Moreover, screening an IL-6-HaloTag®-odorant receptor library with allyl phenyl acetate, revealed both known receptors as best responders for this compound. In summary, the IL-6-HaloTag® represents a promising tool for the de-orphaning of ORs.
Keywords: N-terminal epitope tags; Rho-tag; cAMP assay; flow cytometry.
Conflict of interest statement
Competing interests: The authors have declared that no competing interests exist.
Figures




References
LinkOut - more resources
Full Text Sources
Miscellaneous