

Targeting Glioblastoma Stem Cells through Disruption of the Circadian Clock
- PMID: 31455674
- PMCID: PMC6983300
- DOI: 10.1158/2159-8290.CD-19-0215
Targeting Glioblastoma Stem Cells through Disruption of the Circadian Clock
Abstract
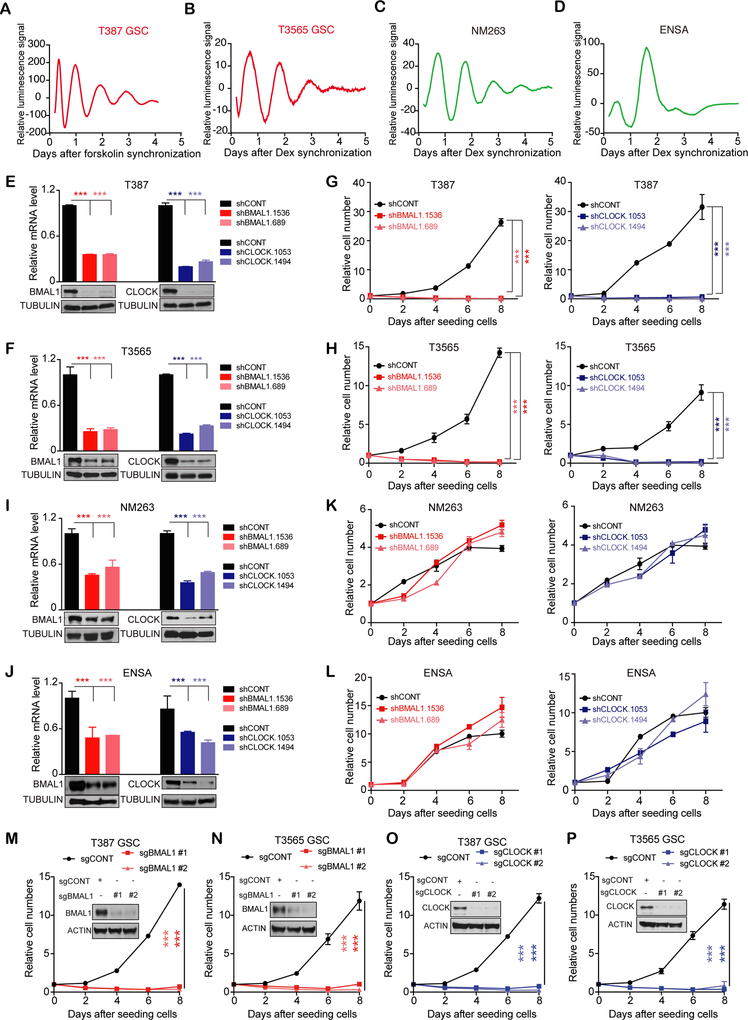
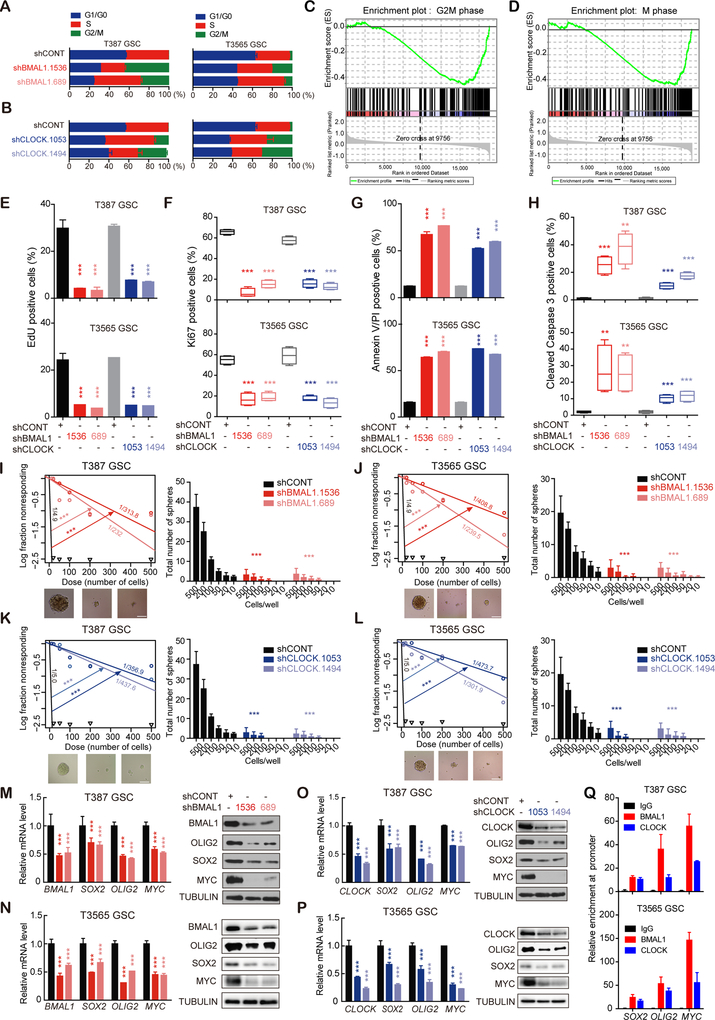
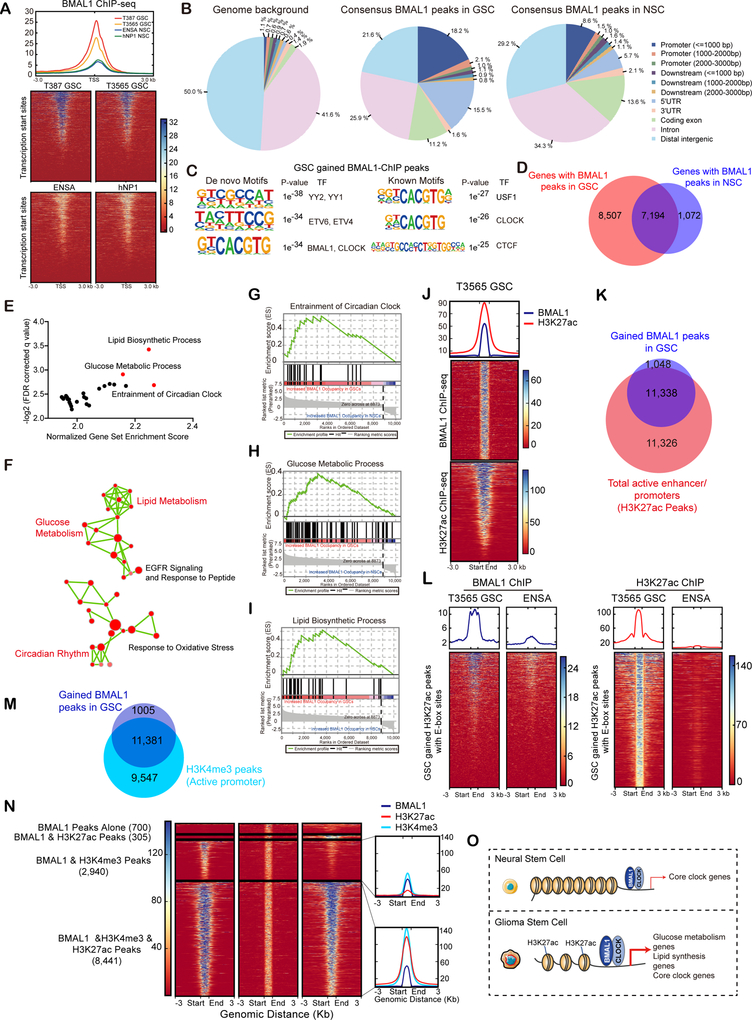
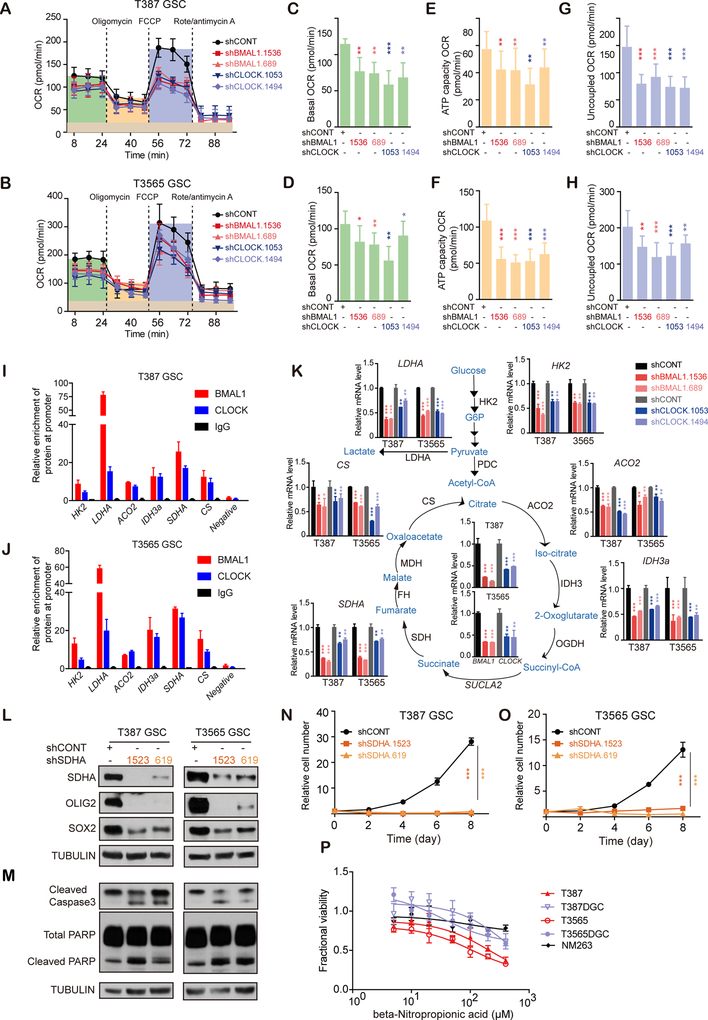
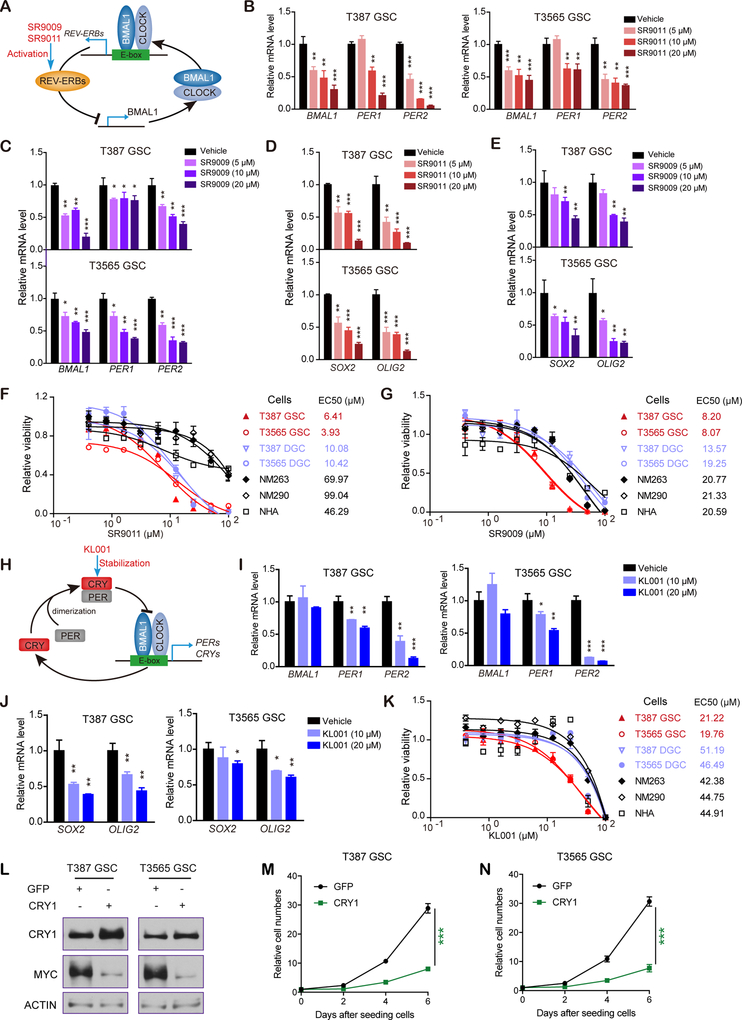
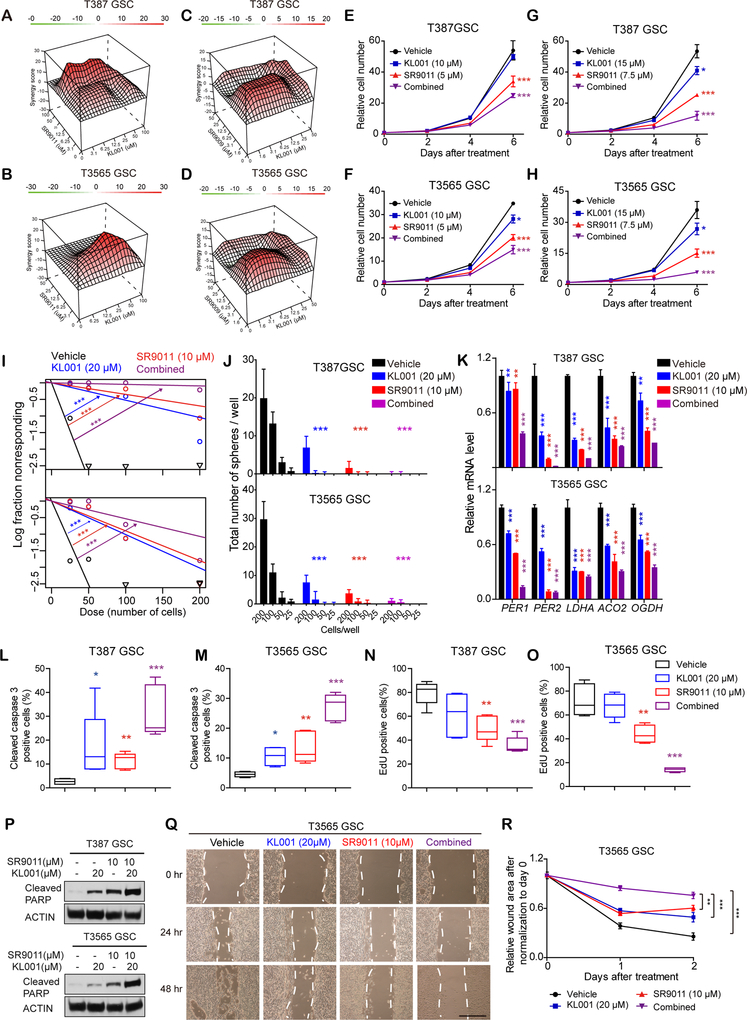
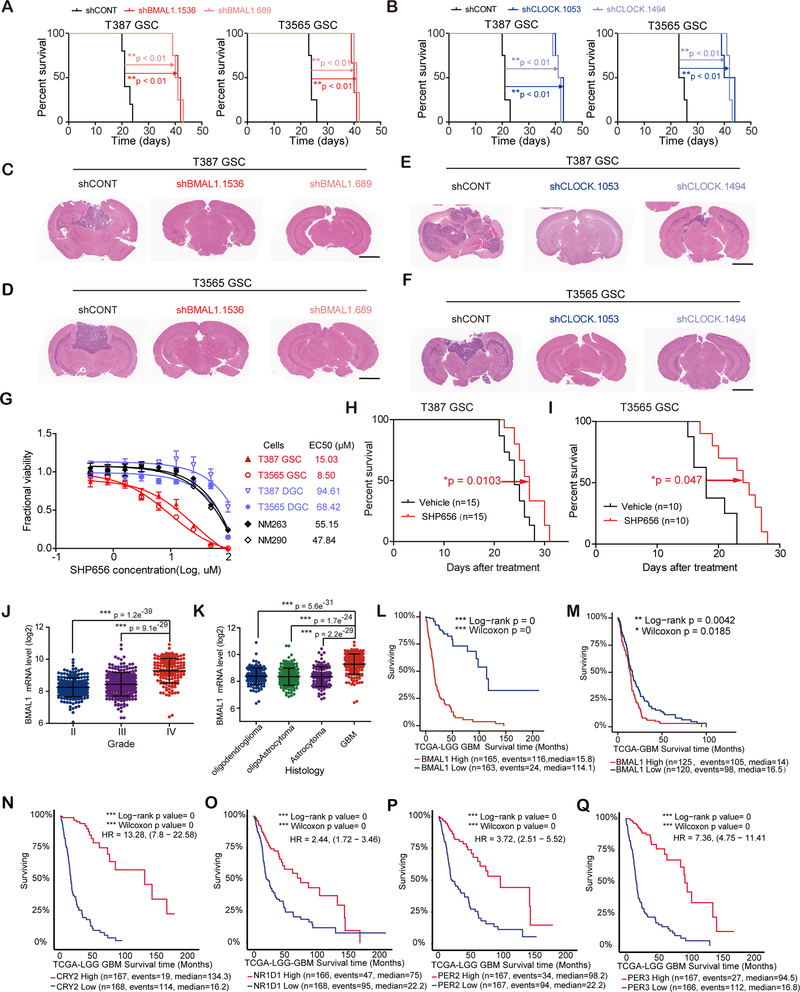
Glioblastomas are highly lethal cancers, containing self-renewing glioblastoma stem cells (GSC). Here, we show that GSCs, differentiated glioblastoma cells (DGC), and nonmalignant brain cultures all displayed robust circadian rhythms, yet GSCs alone displayed exquisite dependence on core clock transcription factors, BMAL1 and CLOCK, for optimal cell growth. Downregulation of BMAL1 or CLOCK in GSCs induced cell-cycle arrest and apoptosis. Chromatin immunoprecipitation revealed that BMAL1 preferentially bound metabolic genes and was associated with active chromatin regions in GSCs compared with neural stem cells. Targeting BMAL1 or CLOCK attenuated mitochondrial metabolic function and reduced expression of tricarboxylic acid cycle enzymes. Small-molecule agonists of two independent BMAL1-CLOCK negative regulators, the cryptochromes and REV-ERBs, downregulated stem cell factors and reduced GSC growth. Combination of cryptochrome and REV-ERB agonists induced synergistic antitumor efficacy. Collectively, these findings show that GSCs co-opt circadian regulators beyond canonical circadian circuitry to promote stemness maintenance and metabolism, offering novel therapeutic paradigms. SIGNIFICANCE: Cancer stem cells are highly malignant tumor-cell populations. We demonstrate that GSCs selectively depend on circadian regulators, with increased binding of the regulators in active chromatin regions promoting tumor metabolism. Supporting clinical relevance, pharmacologic targeting of circadian networks specifically disrupted cancer stem cell growth and self-renewal.This article is highlighted in the In This Issue feature, p. 1469.
©2019 American Association for Cancer Research.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
Comment in
-
Alterations to the circadian clock make brain tumours vulnerable.Nature. 2019 Oct;574(7778):337-338. doi: 10.1038/d41586-019-03042-0. Nature. 2019. PMID: 31611687 No abstract available.
References
-
- Meyer M, Reimand J, Lan X, Head R, Zhu X, Kushida M, et al. Single cell-derived clonal analysis of human glioblastoma links functional and genomic heterogeneity. Proceedings of the National Academy of Sciences of the United States of America 2015;112(3):851–6 doi 10.1073/pnas.1320611111. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 CA169117/CA/NCI NIH HHS/United States
- TL1 TR002549/TR/NCATS NIH HHS/United States
- F30 CA217065/CA/NCI NIH HHS/United States
- F30 CA217066/CA/NCI NIH HHS/United States
- R01 DK108087/DK/NIDDK NIH HHS/United States
- R01 NS087913/NS/NINDS NIH HHS/United States
- R01 NS103434/NS/NINDS NIH HHS/United States
- F30 CA203101/CA/NCI NIH HHS/United States
- R01 NS091080/NS/NINDS NIH HHS/United States
- R01 CA184090/CA/NCI NIH HHS/United States
- R01 NS099175/NS/NINDS NIH HHS/United States
- R01 CA238662/CA/NCI NIH HHS/United States
- R01 CA171652/CA/NCI NIH HHS/United States
- R01 NS089272/NS/NINDS NIH HHS/United States
- F30 CA236313/CA/NCI NIH HHS/United States
- R35 CA197718/CA/NCI NIH HHS/United States
- T32 GM007250/GM/NIGMS NIH HHS/United States
- R01 CA154130/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
