

An Interplay Between Reaction-Diffusion and Cell-Matrix Adhesion Regulates Multiscale Invasion in Early Breast Carcinomatosis
- PMID: 31456688
- PMCID: PMC6700745
- DOI: 10.3389/fphys.2019.00790
An Interplay Between Reaction-Diffusion and Cell-Matrix Adhesion Regulates Multiscale Invasion in Early Breast Carcinomatosis
Abstract
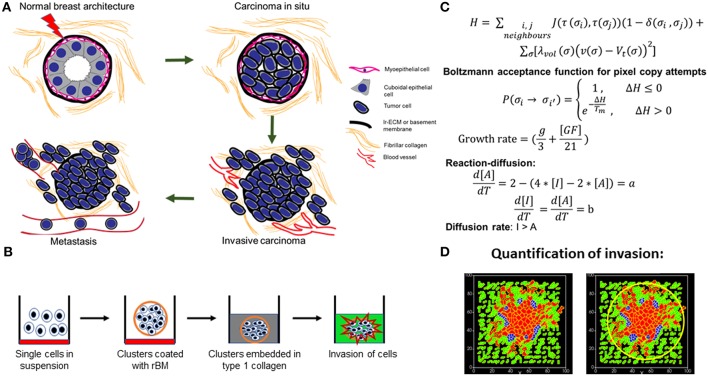
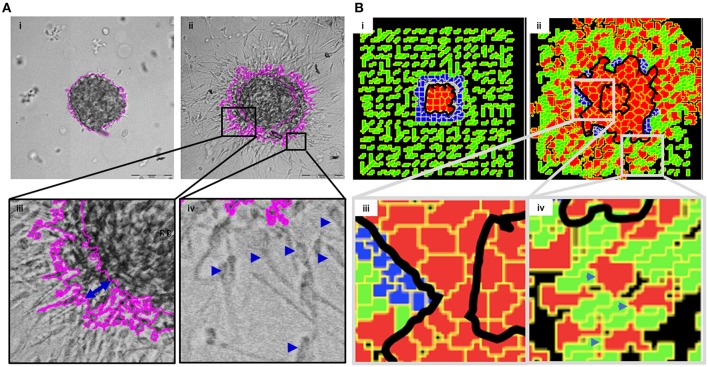
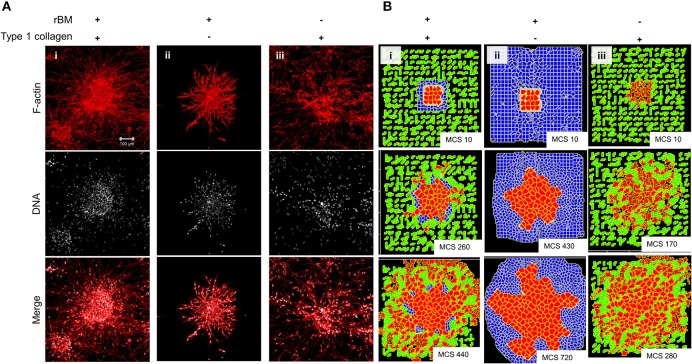
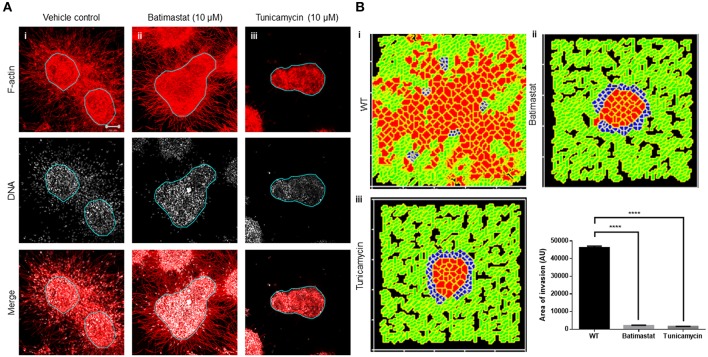
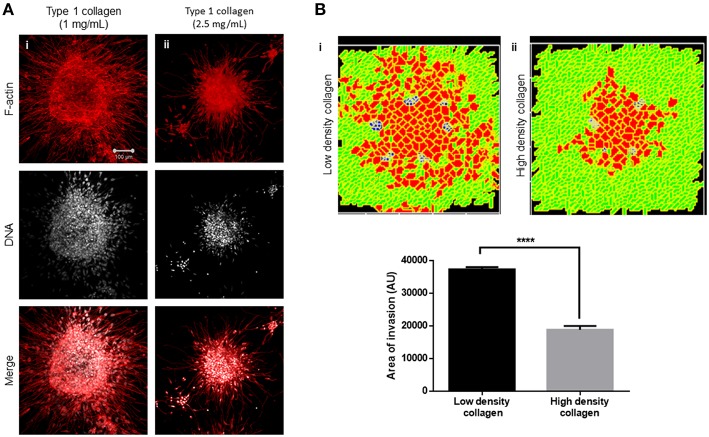
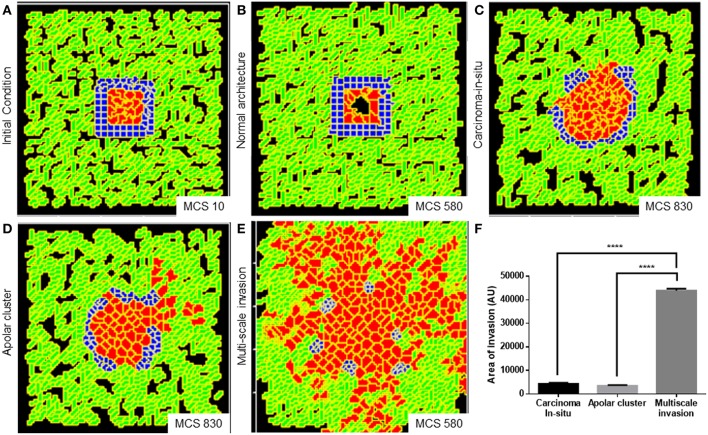
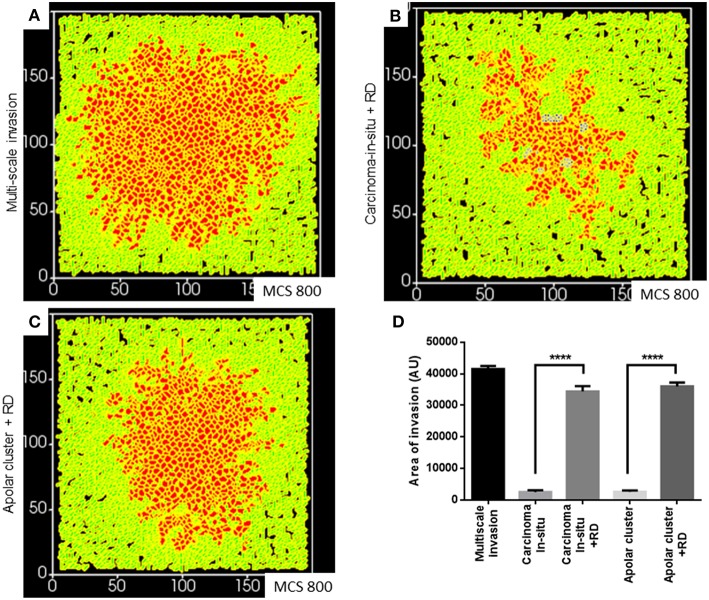
The progression of cancer in the breast involves multiple reciprocal interactions between malignantly transformed epithelia, surrounding untransformed but affected stromal cells, and the extracellular matrix (ECM) that is remodeled during the process. A quantitative understanding of the relative contribution of such interactions to phenotypes associated with cancer cells can be arrived at through the construction of increasingly complex experimental and computational models. Herein, we introduce a multiscale three-dimensional (3D) organo- and pathotypic experimental assay that approximates, to an unprecedented extent, the histopathological complexity of a tumor disseminating into its surrounding stromal milieu via both bulk and solitary motility dynamics. End point and time-lapse microscopic observations of this assay allow us to study the earliest steps of cancer invasion as well as the dynamical interactions between the epithelial and stromal compartments. We then simulate our experimental observations using the modeling environment Compucell3D that is based on the Glazier-Graner-Hogeweg model. The computational model, which comprises adhesion between cancer cells and the matrices, cell proliferation and apoptosis, and matrix remodeling through reaction-diffusion-based morphogen dynamics, is first trained to phenocopy controls run with the experimental model, wherein one or the other matrices have been removed. The trained computational model successfully predicts phenotypes of the experimental counterparts that are subjected to pharmacological treatments (inhibition of N-linked glycosylation and matrix metalloproteinase activity) and scaffold modulation (alteration of collagen density). Further parametric exploration-based simulations suggest that specific permissive regimes of cell-cell and cell-matrix adhesions, operating in the context of a reaction-diffusion-regulated ECM dynamics, promote multiscale invasion of breast cancer cells and determine the extent to which the latter migrate through their surrounding stroma.
Keywords: breast cancer; cell adhesion; cellular potts model (CPM); multiscale invasion; reaction diffusion.
Figures
Similar articles
-
Integrating intracellular dynamics using CompuCell3D and Bionetsolver: applications to multiscale modelling of cancer cell growth and invasion.PLoS One. 2012;7(3):e33726. doi: 10.1371/journal.pone.0033726. Epub 2012 Mar 26. PLoS One. 2012. PMID: 22461894 Free PMC article.
-
Matrix adhesion and remodeling diversifies modes of cancer invasion across spatial scales.J Theor Biol. 2021 Sep 7;524:110733. doi: 10.1016/j.jtbi.2021.110733. Epub 2021 Apr 30. J Theor Biol. 2021. PMID: 33933478
-
FAP-overexpressing fibroblasts produce an extracellular matrix that enhances invasive velocity and directionality of pancreatic cancer cells.BMC Cancer. 2011 Jun 13;11:245. doi: 10.1186/1471-2407-11-245. BMC Cancer. 2011. PMID: 21668992 Free PMC article.
-
The matrix environmental and cell mechanical properties regulate cell migration and contribute to the invasive phenotype of cancer cells.Rep Prog Phys. 2019 Jun;82(6):064602. doi: 10.1088/1361-6633/ab1628. Epub 2019 Apr 4. Rep Prog Phys. 2019. PMID: 30947151 Review.
-
Insidious changes in stromal matrix fuel cancer progression.Mol Cancer Res. 2014 Mar;12(3):297-312. doi: 10.1158/1541-7786.MCR-13-0535. Epub 2014 Jan 22. Mol Cancer Res. 2014. PMID: 24452359 Free PMC article. Review.
Cited by
-
The senescent mesothelial matrix accentuates colonization by ovarian cancer cells.Cell Mol Life Sci. 2023 Dec 3;81(1):2. doi: 10.1007/s00018-023-05017-x. Cell Mol Life Sci. 2023. PMID: 38043093 Free PMC article.
-
An Endosomal Acid-Regulatory Feedback System Rewires Cytosolic cAMP Metabolism and Drives Tumor Progression.Mol Cancer Res. 2024 May 2;22(5):465-481. doi: 10.1158/1541-7786.MCR-23-0606. Mol Cancer Res. 2024. PMID: 38319300 Free PMC article.
-
Breast Cancer: Extracellular Matrix and Microbiome Interactions.Int J Mol Sci. 2024 Jun 30;25(13):7226. doi: 10.3390/ijms25137226. Int J Mol Sci. 2024. PMID: 39000333 Free PMC article. Review.
-
Extracellular matrix remodeling in tumor progression and immune escape: from mechanisms to treatments.Mol Cancer. 2023 Mar 11;22(1):48. doi: 10.1186/s12943-023-01744-8. Mol Cancer. 2023. PMID: 36906534 Free PMC article. Review.
-
Size matters: tissue size as a marker for a transition between reaction-diffusion regimes in spatio-temporal distribution of morphogens.R Soc Open Sci. 2022 Jan 26;9(1):211112. doi: 10.1098/rsos.211112. eCollection 2022 Jan. R Soc Open Sci. 2022. PMID: 35116146 Free PMC article.
References
-
- Bachmeier B. E., Nerlich A. G., Lichtinghagen R., Sommerhoff C. P. (2001). Matrix metalloproteinases (MMPs) in breast cancer cell lines of different tumorigenicity. Anticancer Res. 21, 3821–3828. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources