

The Projection Targets of Medium Spiny Neurons Govern Cocaine-Evoked Synaptic Plasticity in the Nucleus Accumbens
- PMID: 31461643
- PMCID: PMC6733522
- DOI: 10.1016/j.celrep.2019.07.074
The Projection Targets of Medium Spiny Neurons Govern Cocaine-Evoked Synaptic Plasticity in the Nucleus Accumbens
Abstract
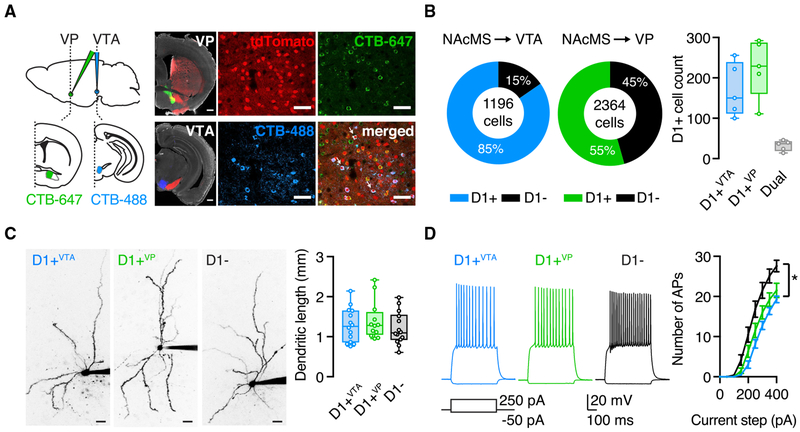
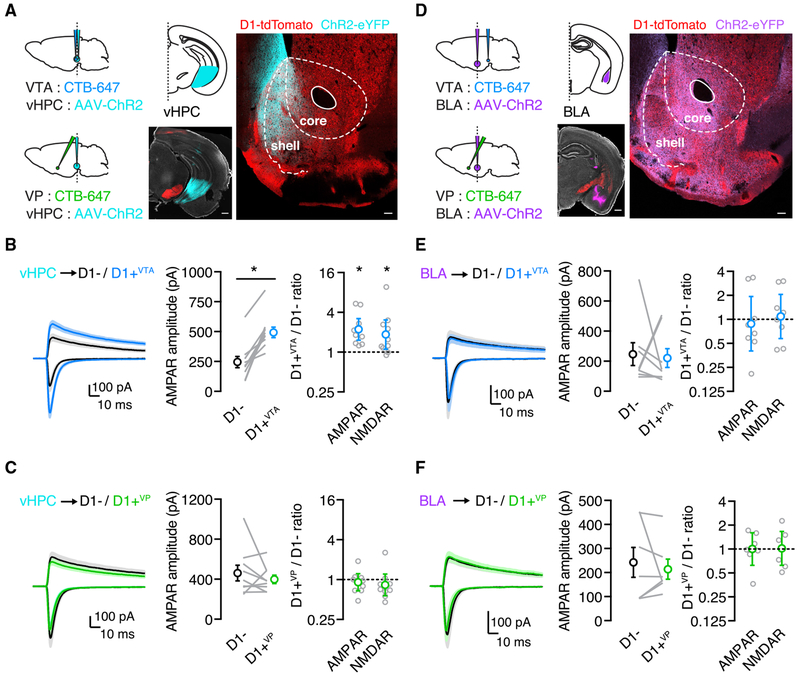
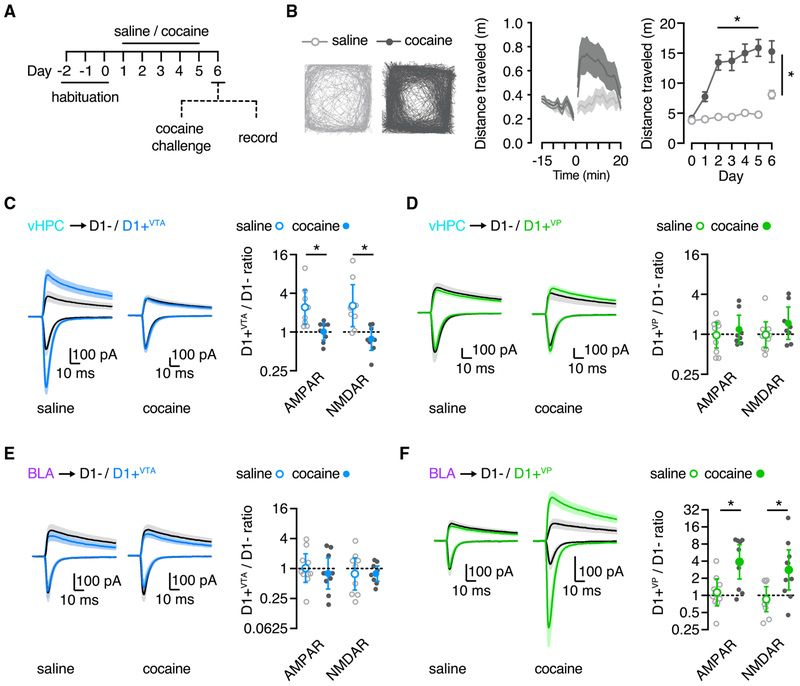
We examine synaptic connectivity and cocaine-evoked plasticity at specific networks within the nucleus accumbens (NAc). We identify distinct subpopulations of D1+ medium spiny neurons (MSNs) that project to either the ventral pallidum (D1+VP) or the ventral tegmental area (D1+VTA). We show that inputs from the ventral hippocampus (vHPC), but not the basolateral amygdala (BLA), are initially biased onto D1+VTA MSNs. However, repeated cocaine exposure eliminates the bias of vHPC inputs onto D1+VTA MSNs, while strengthening BLA inputs onto D1+VP MSNs. Our results reveal that connectivity and plasticity depend on the specific inputs and outputs of D1+ MSNs and highlight the complexity of cocaine-evoked circuit level adaptations in the NAc.
Keywords: cocaine sensitization; medium spiny neuron; nucleus accumbens; spiny projection neuron; synaptic plasticity.
Copyright © 2019 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
The authors declare no competing interests.
Figures
References
-
- Bocklisch C, Pascoli V, Wong JCY, House DRC, Yvon C, de Roo M, Tan KR, and Lüscher C (2013). Cocaine disinhibits dopamine neurons by potentiation of GABA transmission in the ventral tegmental area. Science 341, 1521–1525. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
