

A Bile Duct-on-a-Chip With Organ-Level Functions
- PMID: 31465556
- PMCID: PMC7048662
- DOI: 10.1002/hep.30918
A Bile Duct-on-a-Chip With Organ-Level Functions
Abstract
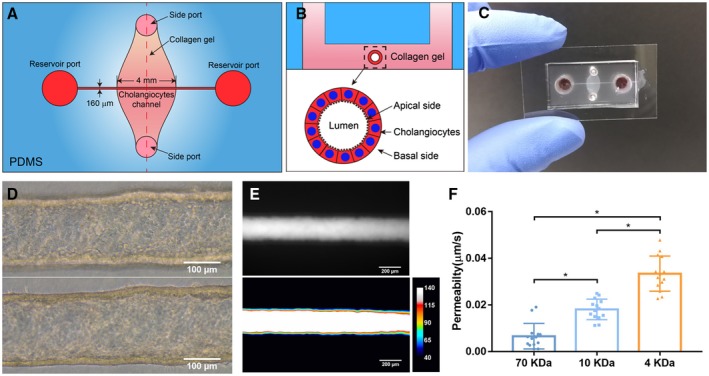
Background and aims: Chronic cholestatic liver diseases, such as primary biliary cholangitis (PBC) and primary sclerosing cholangitis (PSC), are frequently associated with damage to the barrier function of the biliary epithelium. Here, we report on a bile duct-on-a-chip that phenocopies not only the tubular architecture of the bile duct in three dimensions, but also its barrier functions.
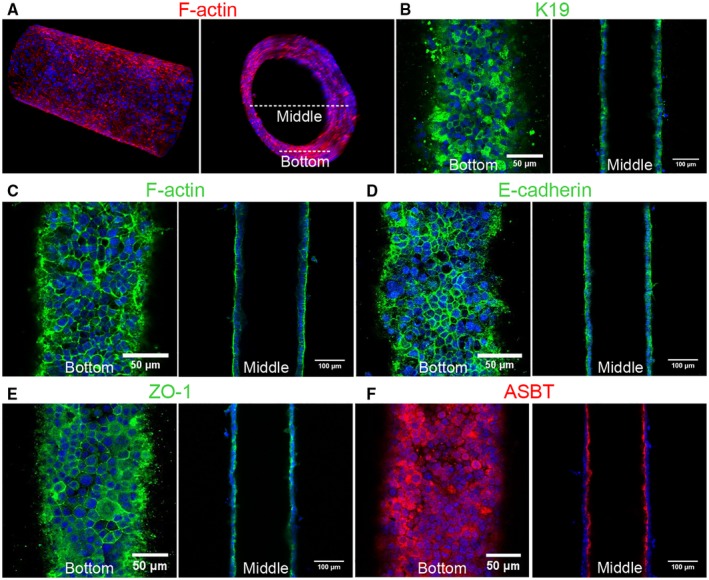
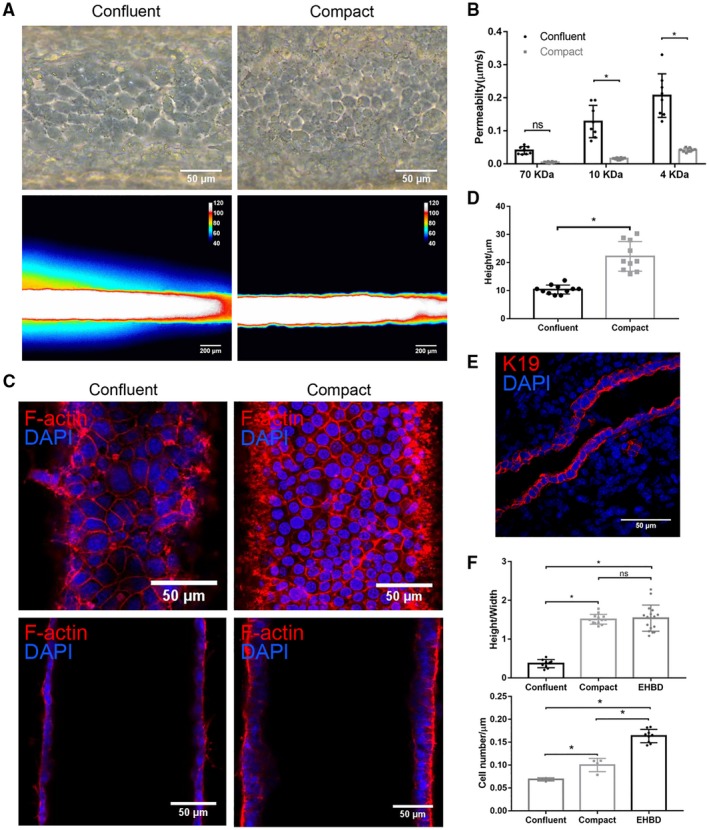
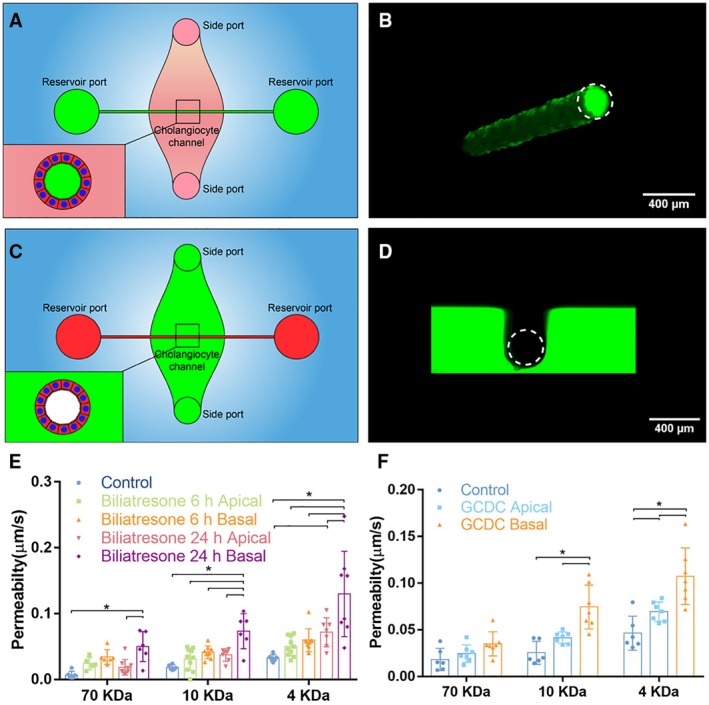
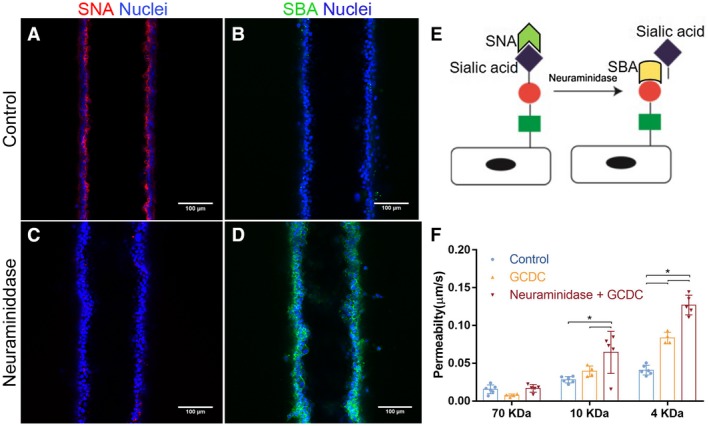
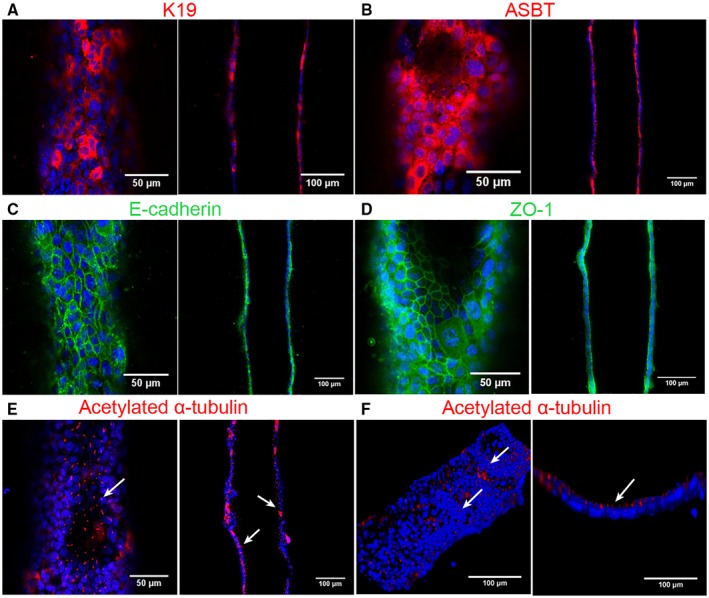
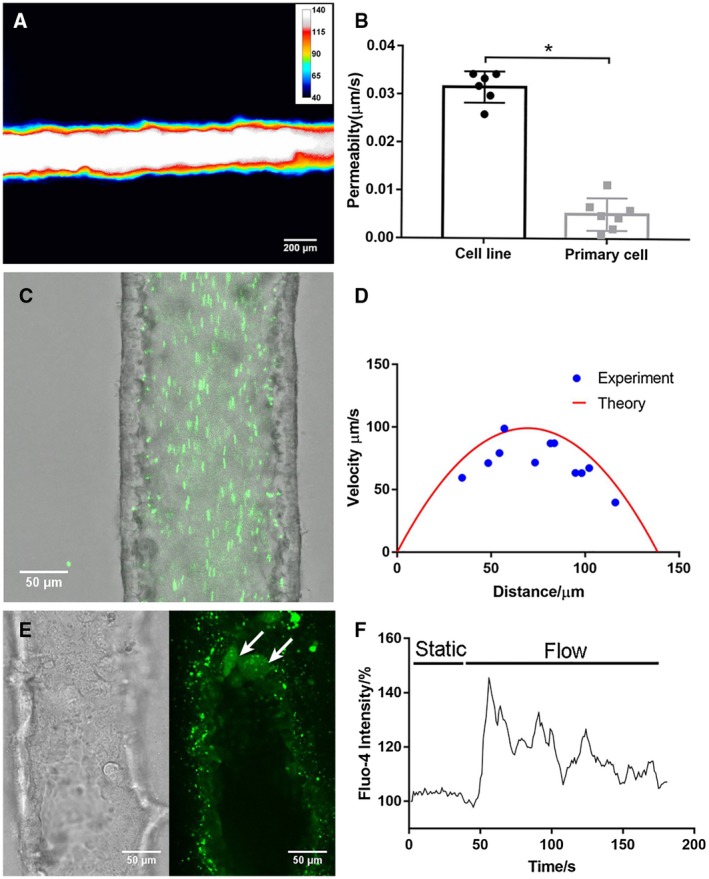
Approach and results: We showed that mouse cholangiocytes in the channel of the device became polarized and formed mature tight junctions, that the permeability of the cholangiocyte monolayer was comparable to ex vivo measurements, and that cholangiocytes in the device were mechanosensitive (as demonstrated by changes in calcium flux under applied luminal flow). Permeability decreased significantly when cells formed a compact monolayer with cell densities comparable to those observed in vivo. This device enabled independent access to the apical and basolateral surfaces of the cholangiocyte channel, allowing proof-of-concept toxicity studies with the biliary toxin, biliatresone, and the bile acid, glycochenodeoxycholic acid. The cholangiocyte basolateral side was more vulnerable than the apical side to treatment with either agent, suggesting a protective adaptation of the apical surface that is normally exposed to bile. Further studies revealed a protective role of the cholangiocyte apical glycocalyx, wherein disruption of the glycocalyx with neuraminidase increased the permeability of the cholangiocyte monolayer after treatment with glycochenodeoxycholic acid.
Conclusions: This bile duct-on-a-chip captured essential features of a simplified bile duct in structure and organ-level functions and represents an in vitro platform to study the pathophysiology of the bile duct using cholangiocytes from a variety of sources.
© 2019 The Authors. Hepatology published by Wiley Periodicals, Inc., on behalf of American Association for the Study of Liver Diseases.
Figures
Comment in
-
Letter to the Editor: Comment on "A Bile Duct-on-a-Chip With Organ-Level Functions".Hepatology. 2021 Feb;73(2):872. doi: 10.1002/hep.31470. Hepatology. 2021. PMID: 32687628 No abstract available.
-
REPLY.Hepatology. 2021 Feb;73(2):872-873. doi: 10.1002/hep.31471. Hepatology. 2021. PMID: 33128466 No abstract available.
References
-
- Matsumoto K, Imasato M, Yamazaki Y, Tanaka H, Watanabe M, Eguchi H, Nagano H, et al. Claudin 2 deficiency reduces bile flow and increases susceptibility to cholesterol gallstone disease in mice. Gastroenterology 2014;147:1134‐1145.e10. - PubMed
-
- Sakisaka S, Kawaguchi T, Taniguchi E, Hanada S, Sasatomi K, Koga H, et al. Alterations in tight junctions differ between primary biliary cirrhosis and primary sclerosing cholangitis. Hepatology 2001;33:1460‐1468. - PubMed
-
- Nakanuma Y, Tsuneyama K, Gershwin ME, Yasoshima M. Pathology and immunopathology of primary biliary cirrhosis with emphasis on bile duct lesions: recent progress. Semin Liver Dis 1995;15:313‐328. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
