

Opioid and chemokine regulation of cortical synaptodendritic damage in HIV-associated neurocognitive disorders
- PMID: 31465771
- PMCID: PMC6766413
- DOI: 10.1016/j.brainres.2019.146409
Opioid and chemokine regulation of cortical synaptodendritic damage in HIV-associated neurocognitive disorders
Abstract
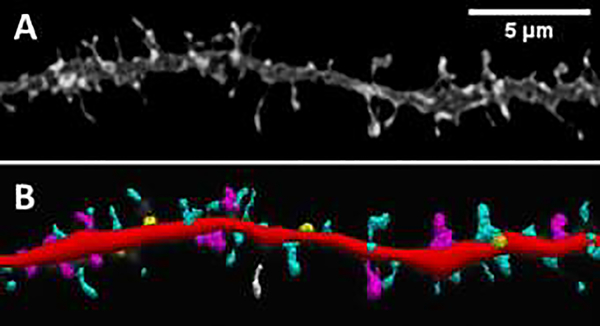
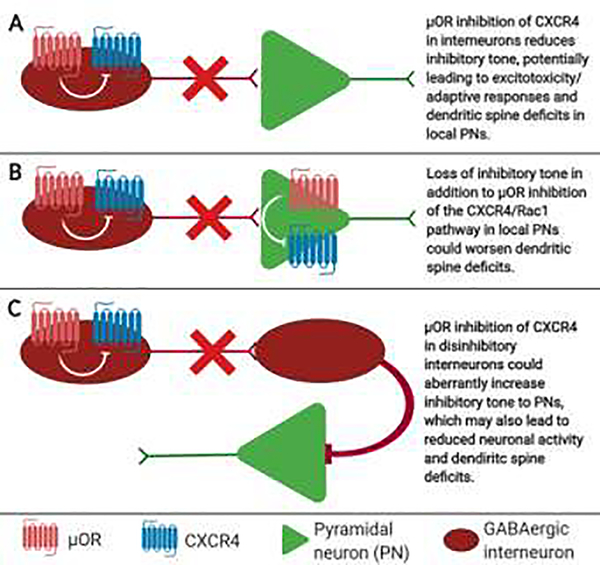
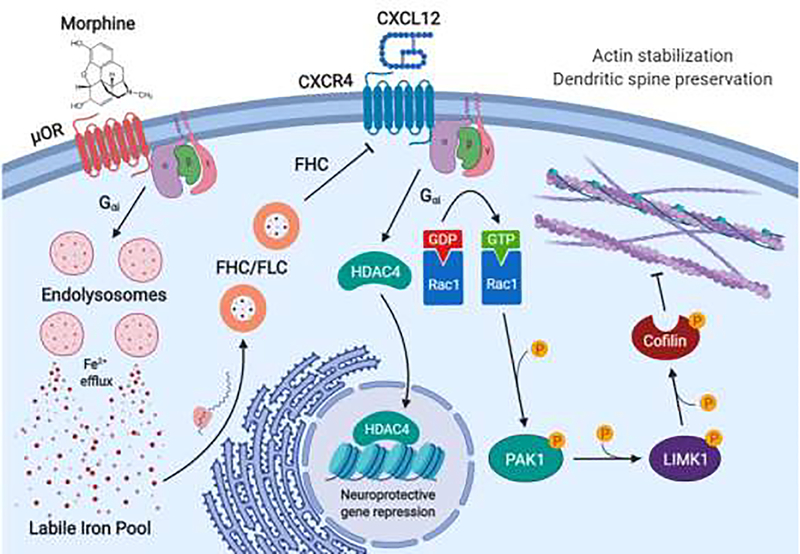
Human immunodeficiency virus (HIV)-associated neurocognitive disorders (HAND) persist despite effective antiretroviral therapies (ART). Evidence suggests that modern HAND is driven by subtle synaptodendritic damage in select brain regions, as ART-treated patients do not display overt neuronal death in postmortem brain studies. HAND symptoms are also aggravated by drug abuse, particularly with injection opioids. Opioid use produces region-specific synaptodendritic damage in similar brain regions, suggesting a convergent mechanism that may enhance HAND progression in opioid-using patients. Importantly, studies indicate that synaptodendritic damage and cognitive impairment in HAND may be reversible. Activation of the homeostatic chemokine receptor CXCR4 by its natural ligand CXCL12 positively regulates neuronal survival and dendritic spine density in cortical neurons, reducing functional deficits. However, the molecular mechanisms that underlie CXCR4, as well as opioid-mediated regulation of dendritic spines are not completely defined. Here, we will consolidate studies that describe the region-specific synaptodendritic damage in the cerebral cortex of patients and animal models of HAND, describe the pathways by which opioids may contribute to cortical synaptodendritic damage, and discuss the prospects of using the CXCR4 signaling pathway to identify new approaches to reverse dendritic spine deficits. Additionally, we will discuss novel research questions that have emerged from recent studies of CXCR4 and µ-opioid actions in the cortex. Understanding the pathways that underlie synaptodendritic damage and rescue are necessary for developing novel, effective therapeutics for this growing patient population.
Keywords: CXCL12; CXCR4; Dendritic spines; HAND; Opioids; neuroHIV.
Copyright © 2019. Published by Elsevier B.V.
Figures

Similar articles
-
Opioid Modulation of Neuronal Iron and Potential Contributions to NeuroHIV.Methods Mol Biol. 2021;2201:139-162. doi: 10.1007/978-1-0716-0884-5_13. Methods Mol Biol. 2021. PMID: 32975796 Free PMC article. Review.
-
CXCL12-induced rescue of cortical dendritic spines and cognitive flexibility.Elife. 2020 Jan 23;9:e49717. doi: 10.7554/eLife.49717. Elife. 2020. PMID: 31971513 Free PMC article.
-
CXCL12 Engages Cortical Inhibitory Neurons to Enhance Dendritic Spine Plasticity and Structured Network Activity.J Neurosci. 2025 Jun 11;45(24):e2213242025. doi: 10.1523/JNEUROSCI.2213-24.2025. J Neurosci. 2025. PMID: 40335156 Free PMC article.
-
Mechanisms of neuronal dysfunction in HIV-associated neurocognitive disorders.Cell Mol Life Sci. 2021 May;78(9):4283-4303. doi: 10.1007/s00018-021-03785-y. Epub 2021 Feb 13. Cell Mol Life Sci. 2021. PMID: 33585975 Free PMC article. Review.
-
Neuronal ferritin heavy chain and drug abuse affect HIV-associated cognitive dysfunction.J Clin Invest. 2014 Feb;124(2):656-69. doi: 10.1172/JCI70090. Epub 2014 Jan 9. J Clin Invest. 2014. PMID: 24401274 Free PMC article.
Cited by
-
The single-cell opioid responses in the context of HIV (SCORCH) consortium.Mol Psychiatry. 2024 Dec;29(12):3950-3961. doi: 10.1038/s41380-024-02620-7. Epub 2024 Jun 15. Mol Psychiatry. 2024. PMID: 38879719 Free PMC article.
-
Accelerated brain aging with opioid misuse and HIV: New insights on the role of glially derived pro-inflammation mediators and neuronal chloride homeostasis.Curr Opin Neurobiol. 2023 Feb;78:102653. doi: 10.1016/j.conb.2022.102653. Epub 2022 Dec 28. Curr Opin Neurobiol. 2023. PMID: 36584655 Free PMC article. Review.
-
Increasing Neuroinflammation Relates to Increasing Neurodegeneration in People with HIV.Viruses. 2023 Aug 30;15(9):1835. doi: 10.3390/v15091835. Viruses. 2023. PMID: 37766242 Free PMC article.
-
Advances in the Experimental Models of HIV-Associated Neurological Disorders.Curr HIV/AIDS Rep. 2021 Oct;18(5):459-474. doi: 10.1007/s11904-021-00570-1. Epub 2021 Aug 24. Curr HIV/AIDS Rep. 2021. PMID: 34427869 Free PMC article. Review.
-
Cellular, synaptic, and network effects of chemokines in the central nervous system and their implications to behavior.Pharmacol Rep. 2021 Dec;73(6):1595-1625. doi: 10.1007/s43440-021-00323-2. Epub 2021 Aug 26. Pharmacol Rep. 2021. PMID: 34498203 Free PMC article. Review.
References
-
- Arber S, Barbayannis FA, Hanser H, Schneider C, Stanyon CA, Bernard O, Caroni P, 1998. Regulation of actin dynamics through phosphorylation of cofilin by LIM-kinase. Nature. 393, 805–9. - PubMed
-
- Arosio P, Carmona F, Gozzelino R, Maccarinelli F, Poli M, 2015. The importance of eukaryotic ferritins in iron handling and cytoprotection. Biochem J. 472, 1–15. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
