

Gasdermin D in peripheral myeloid cells drives neuroinflammation in experimental autoimmune encephalomyelitis
- PMID: 31467036
- PMCID: PMC6829591
- DOI: 10.1084/jem.20190377
Gasdermin D in peripheral myeloid cells drives neuroinflammation in experimental autoimmune encephalomyelitis
Abstract
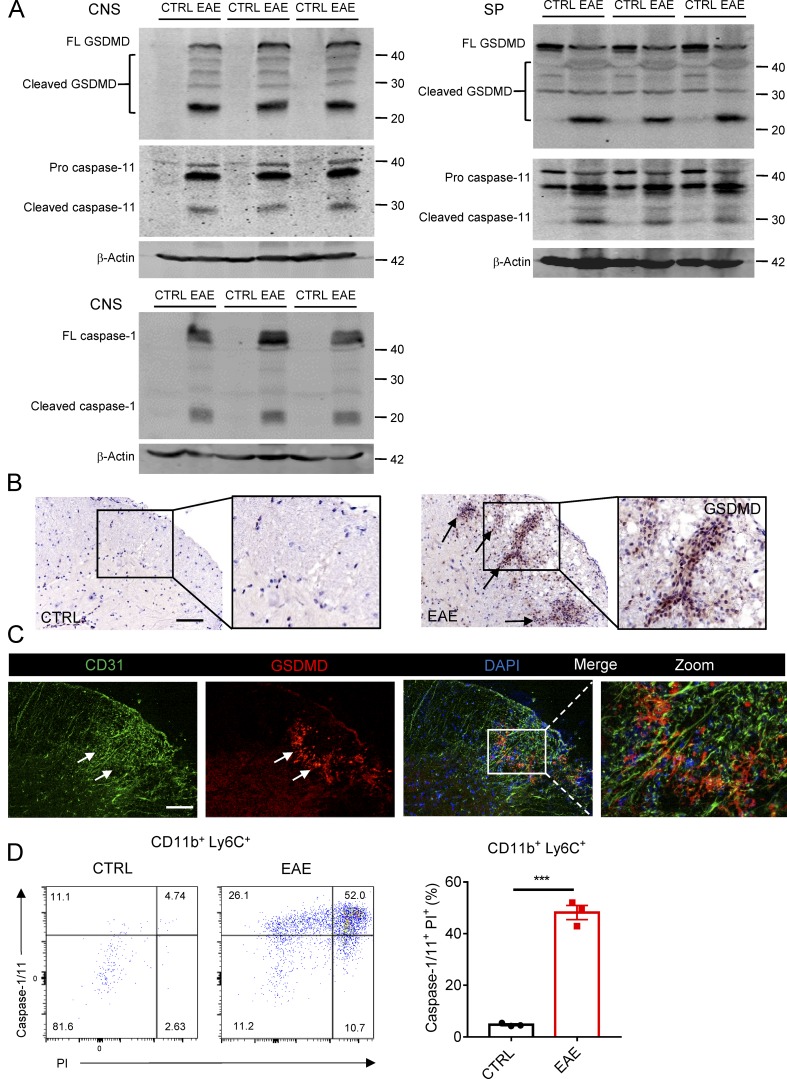
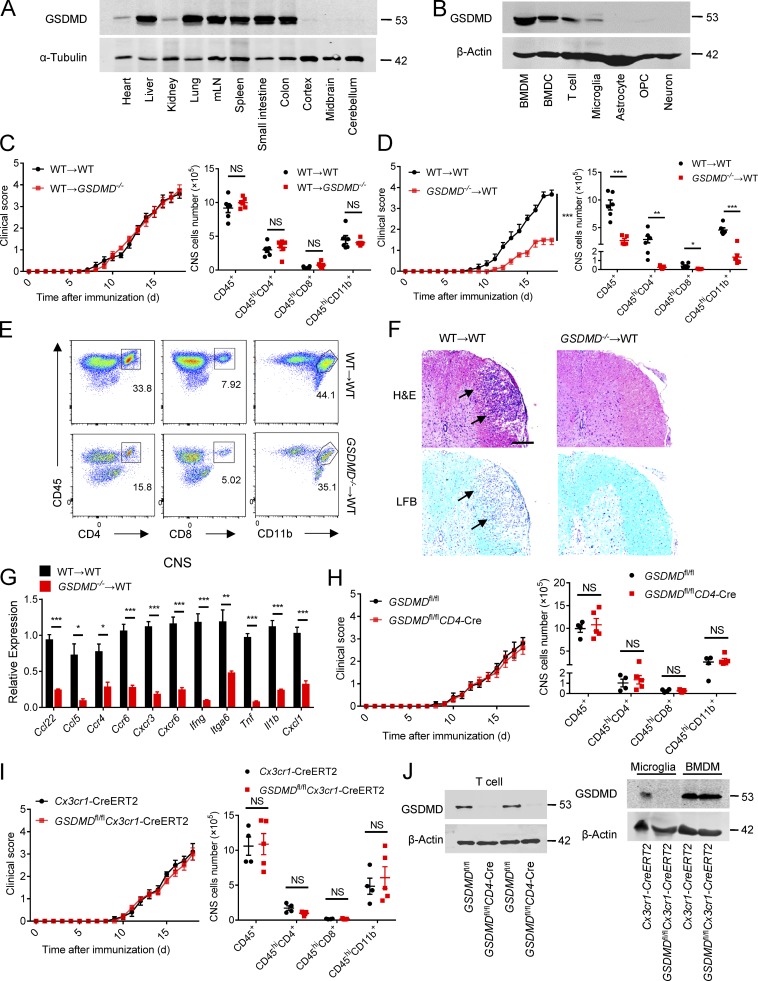
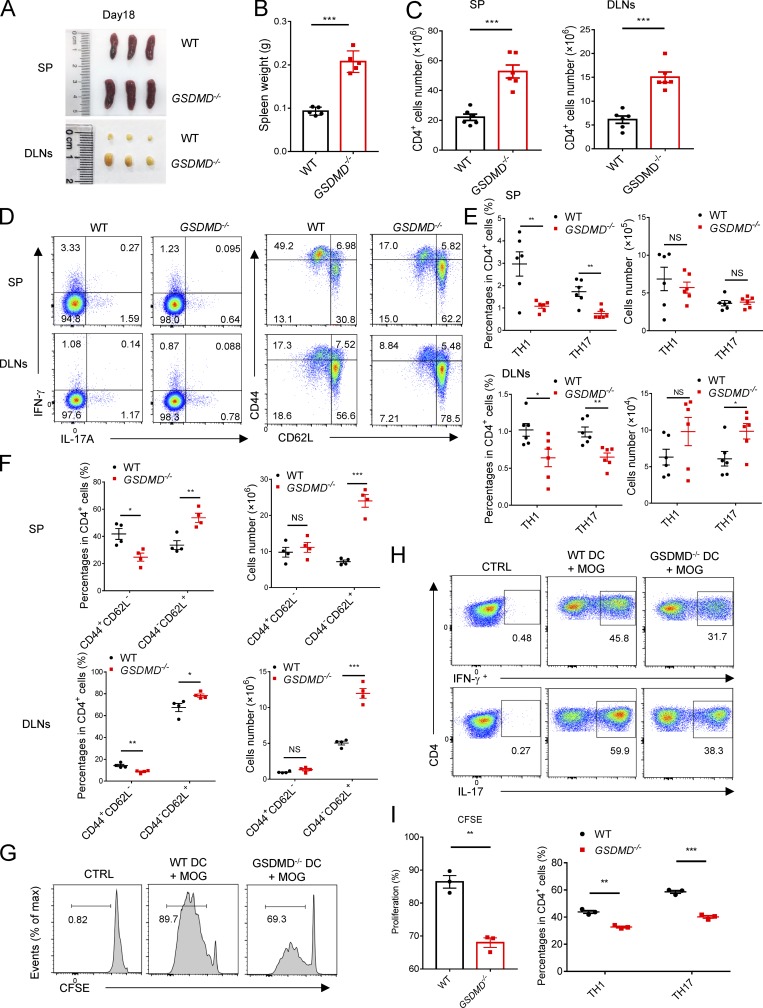
The NLRP3 inflammasome is critical for EAE pathogenesis; however, the role of gasdermin D (GSDMD), a newly identified pyroptosis executioner downstream of NLRP3 inflammasome, in EAE has not been well defined. Here, we observed that the levels of GSDMD protein were greatly enhanced in the CNS of EAE mice, especially near the areas surrounding blood vessels. GSDMD was required for the pathogenesis of EAE, and GSDMD deficiency in peripheral myeloid cells impaired the infiltration of immune cells into the CNS, leading to the suppression of neuroinflammation and demyelination. Furthermore, the loss of GSDMD reduced the activation and differentiation of T cell in the secondary lymphoid organs and prevented T cell infiltration into CNS of EAE. The administration of inflammasome-related cytokines partially rescued the impairment of pathogenesis of EAE in GSDMD KO mice. Collectively, these findings provide the first demonstration of GSDMD in peripheral myeloid cells driving neuroinflammation during EAE pathogenesis.
© 2019 Crown copyright. The government of Australia, Canada, or the UK ("the Crown") owns the copyright interests of authors who are government employees. The Crown Copyright is not transferable.
Figures








References
-
- Banerjee I., Behl B., Mendonca M., Shrivastava G., Russo A.J., Menoret A., Ghosh A., Vella A.T., Vanaja S.K., Sarkar S.N., et al. . 2018. Gasdermin D Restrains Type I Interferon Response to Cytosolic DNA by Disrupting Ionic Homeostasis. Immunity. 49:413–426.e5. 10.1016/j.immuni.2018.07.006 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
