

The rhg1-a (Rhg1 low-copy) nematode resistance source harbors a copia-family retrotransposon within the Rhg1- encoded α-SNAP gene
- PMID: 31468029
- PMCID: PMC6712407
- DOI: 10.1002/pld3.164
The rhg1-a (Rhg1 low-copy) nematode resistance source harbors a copia-family retrotransposon within the Rhg1- encoded α-SNAP gene
Abstract
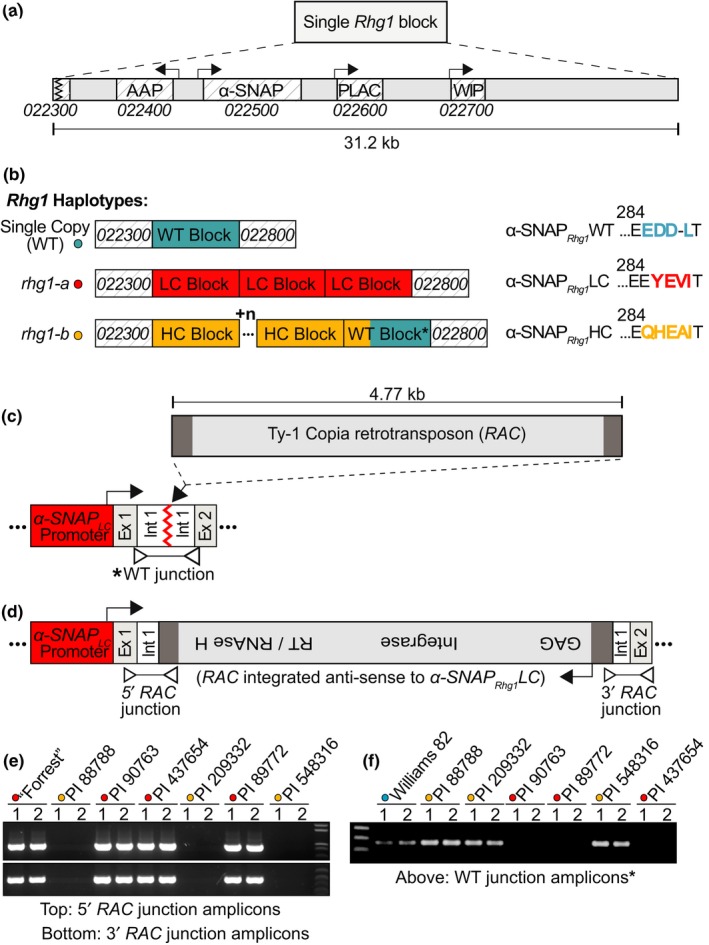
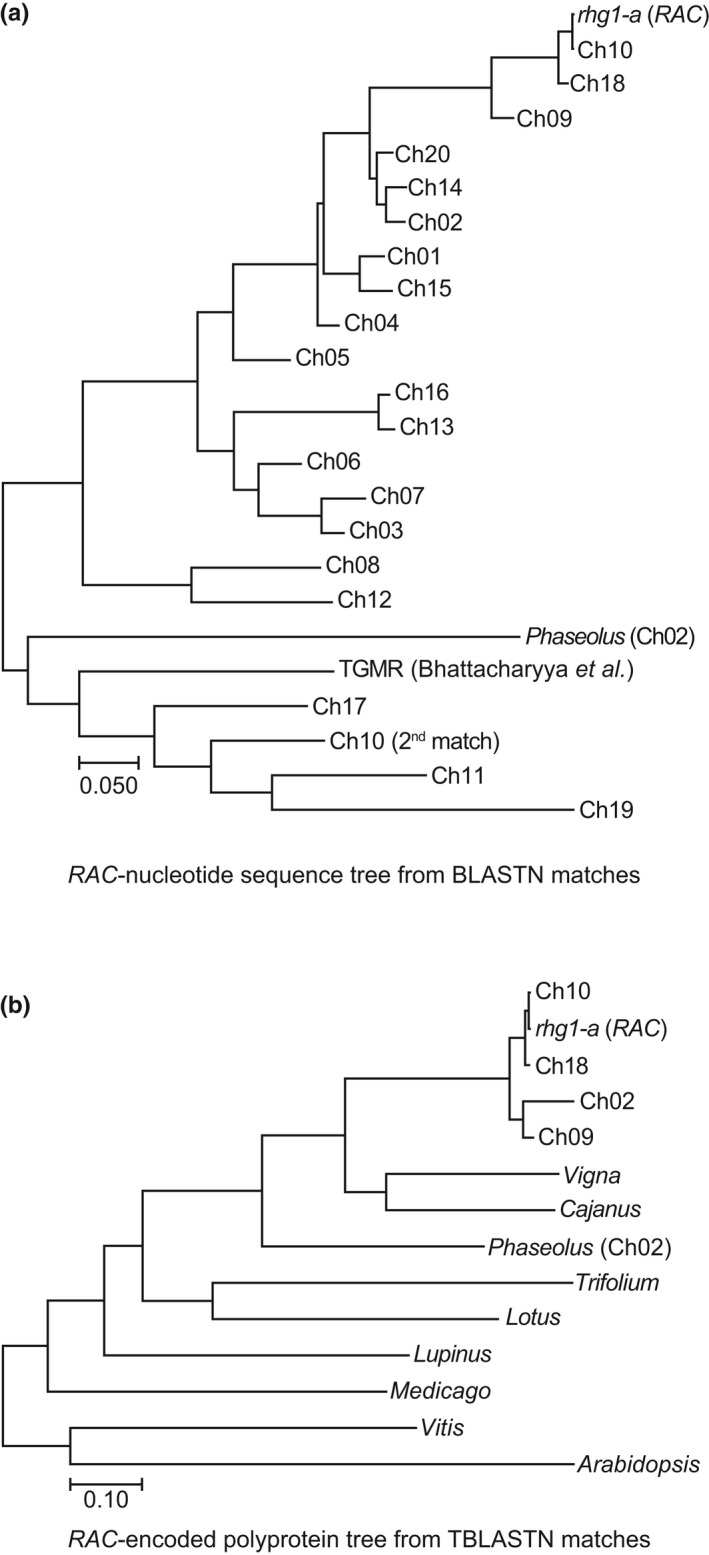
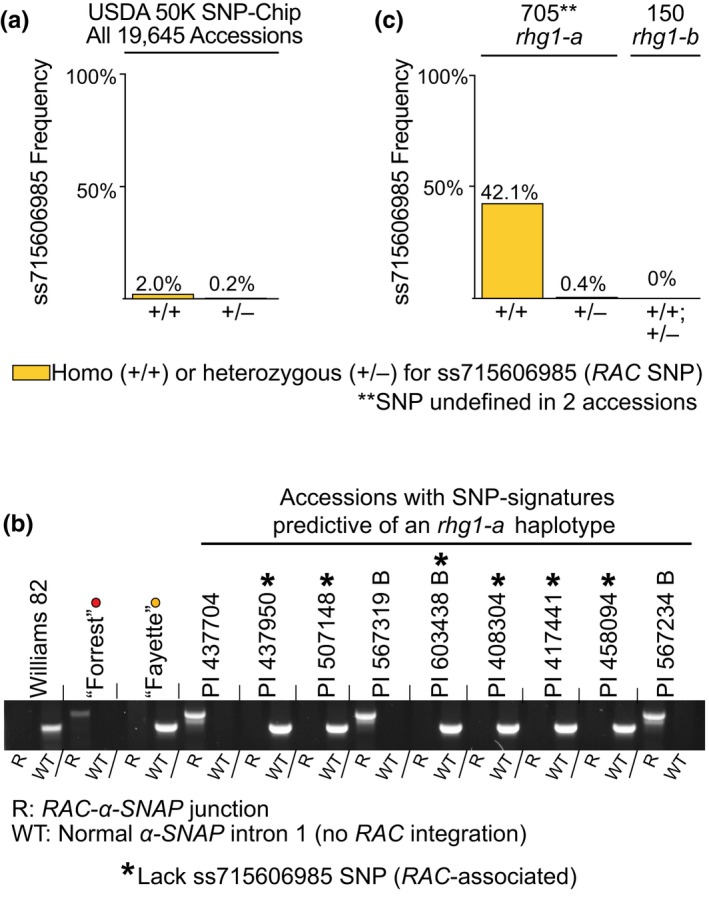
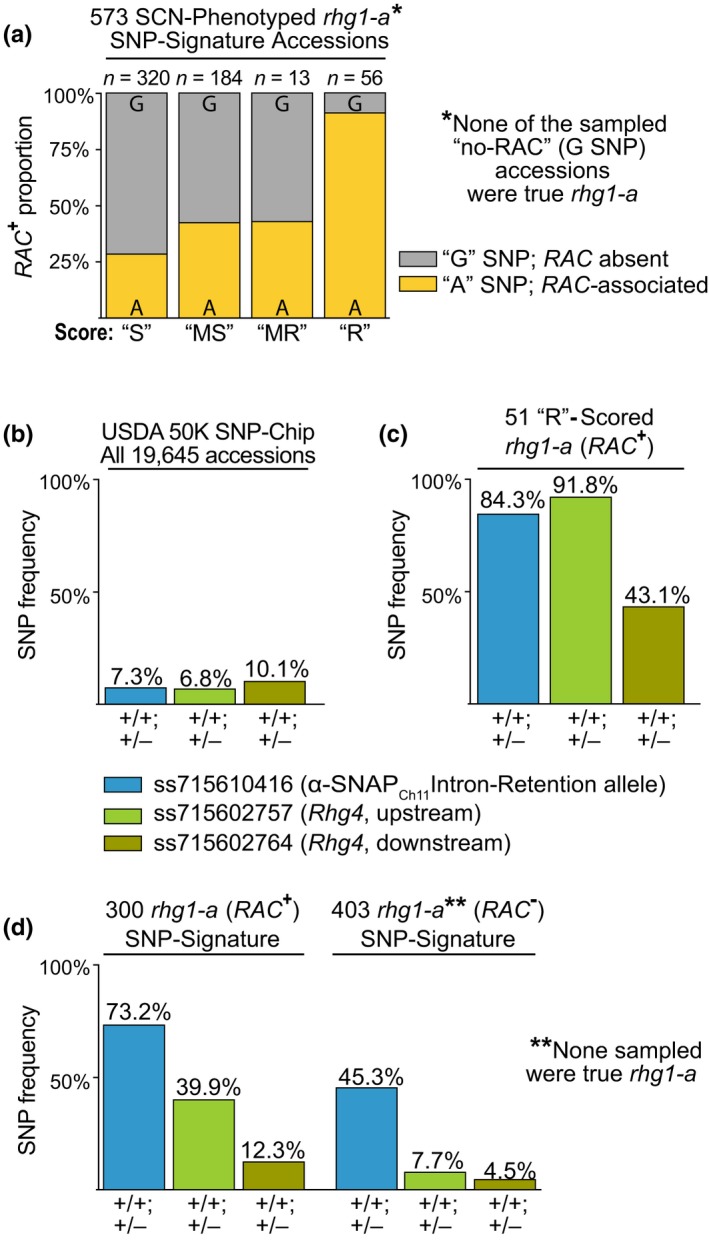
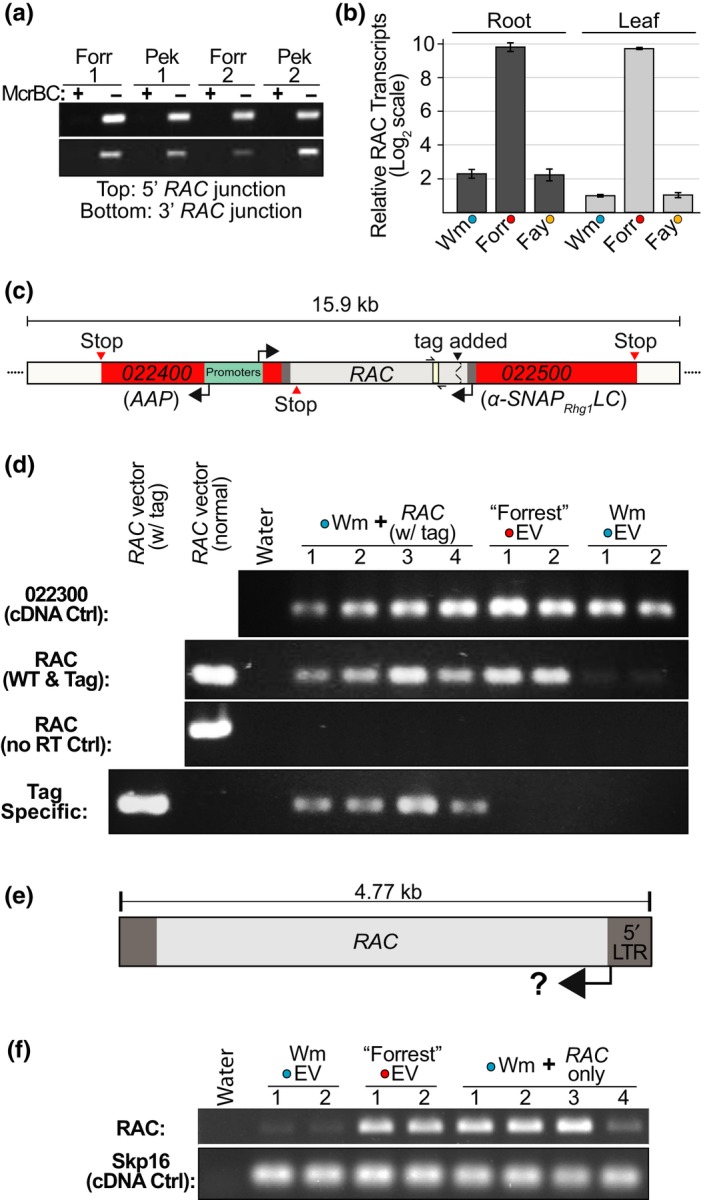
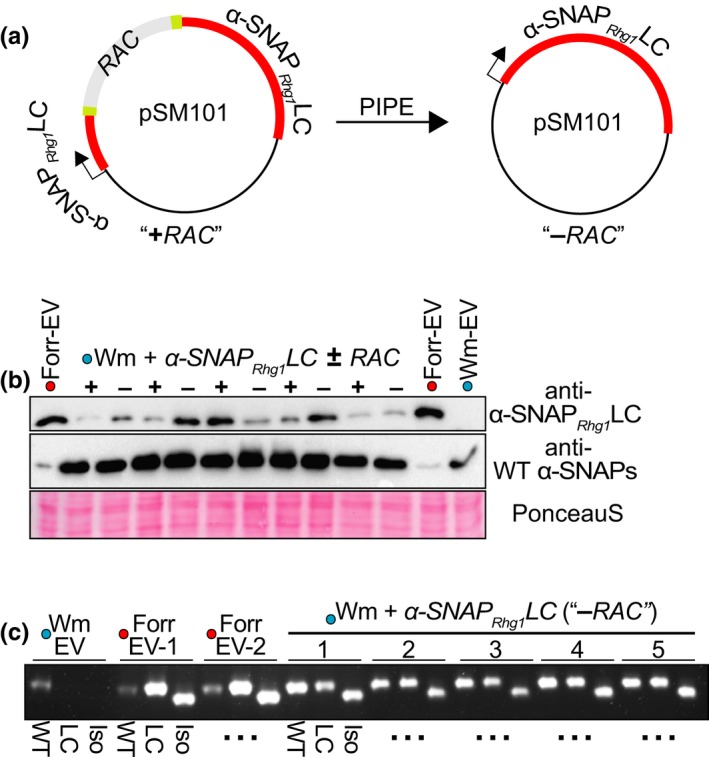
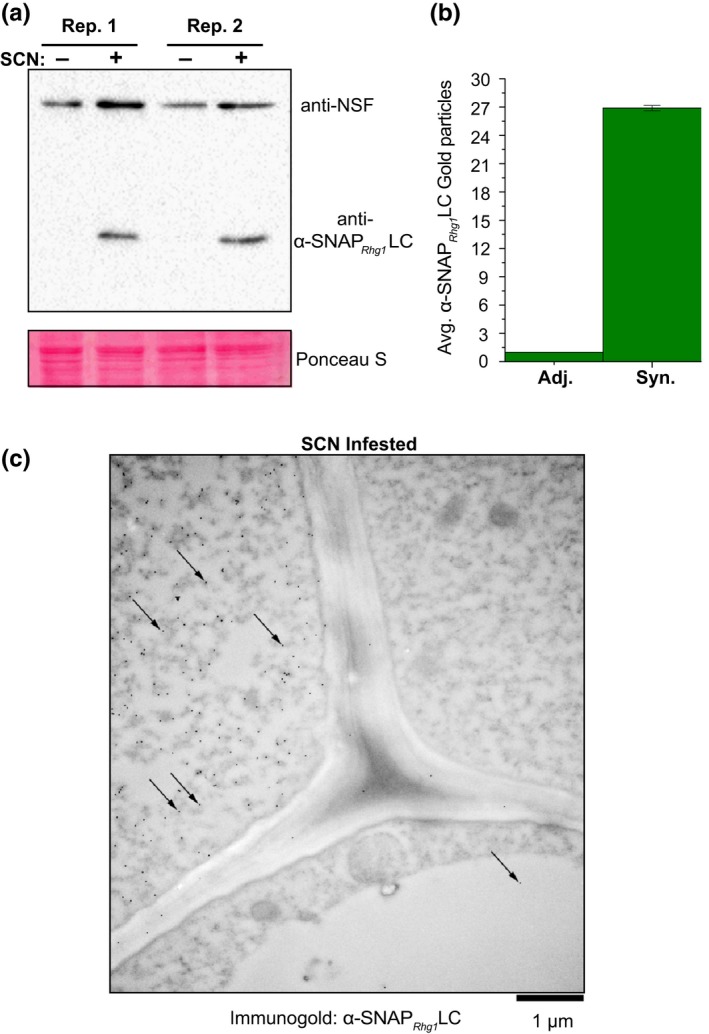
Soybean growers widely use the Resistance to Heterodera glycines 1 (Rhg1) locus to reduce yield losses caused by soybean cyst nematode (SCN). Rhg1 is a tandemly repeated four gene block. Two classes of SCN resistance-conferring Rhg1 haplotypes are recognized: rhg1-a ("Peking-type," low-copy number, three or fewer Rhg1 repeats) and rhg1-b ("PI 88788-type," high-copy number, four or more Rhg1 repeats). The rhg1-a and rhg1-b haplotypes encode α-SNAP (alpha-Soluble NSF Attachment Protein) variants α-SNAP Rhg1 LC and α-SNAP Rhg1 HC, respectively, with differing atypical C-terminal domains, that contribute to SCN resistance. Here we report that rhg1-a soybean accessions harbor a copia retrotransposon within their Rhg1 Glyma.18G022500 (α-SNAP-encoding) gene. We termed this retrotransposon "RAC," for Rhg1 alpha-SNAP copia. Soybean carries multiple RAC-like retrotransposon sequences. The Rhg1 RAC insertion is in the Glyma.18G022500 genes of all true rhg1-a haplotypes we tested and was not detected in any examined rhg1-b or Rhg1WT (single-copy) soybeans. RAC is an intact element residing within intron 1, anti-sense to the rhg1-a α-SNAP open reading frame. RAC has intrinsic promoter activities, but overt impacts of RAC on transgenic α-SNAP Rhg1 LC mRNA and protein abundance were not detected. From the native rhg1-a RAC+ genomic context, elevated α-SNAP Rhg1 LC protein abundance was observed in syncytium cells, as was previously observed for α-SNAP Rhg1 HC (whose rhg1-b does not carry RAC). Using a SoySNP50K SNP corresponding with RAC presence, just ~42% of USDA accessions bearing previously identified rhg1-a SoySNP50K SNP signatures harbor the RAC insertion. Subsequent analysis of several of these putative rhg1-a accessions lacking RAC revealed that none encoded α-SNAPRhg1LC, and thus, they are not rhg1-a. rhg1-a haplotypes are of rising interest, with Rhg4, for combating SCN populations that exhibit increased virulence against the widely used rhg1-b resistance. The present study reveals another unexpected structural feature of many Rhg1 loci, and a selectable feature that is predictive of rhg1-a haplotypes.
Keywords: Rhg1; plant disease resistance; retrotransposon; soybean cyst nematode.
Conflict of interest statement
The authors declare no conflict of interest associated with the work described in this manuscript.
Figures
References
-
- Allen, T. W. , Bradley, C. A. , Sisson, A. J. , Byamukama, E. , Chilvers, M. I. , Coker, C. M. , … Wrather, J. A. (2017). Soybean yield loss estimates due to diseases in the United States and Ontario, Canada, from 2010 to 2014. Plant Health Progress, 18(1), 19–27. 10.1094/PHP-RS-16-0066 - DOI
-
- Anand, S. C. , & Gallo, K. M. (1984) Identification of additional soybean germ plasm with resistance to race 3 of the soybean cyst nematode. Plant Disease, 68(7), 593– 10.1094/PD-69-593 - DOI
-
- Arelli, P. R. , Sleper, D. A. , Yue, P. , & Wilcox, J. A. (2000). Soybean reaction to races 1 and 2 of Heterodera glycines. Contribution of the Missouri Agric. Exp. Stn. Journal Series No. 12,751. Crop Science, 40, 824–826. 10.2135/cropsci2000.403824x - DOI
-
- Bayless, A. M. , Smith, J. M. , Song, J. , McMinn, P. H. , Teillet, A. , August, B. K. , & Bent, A. F. (2016). Disease resistance through impairment of alpha‐SNAP‐NSF interaction and vesicular trafficking by soybean Rhg1 . Proceedings of the National Academy of Sciences of the United States of America, 113, E7375–E7382. - PMC - PubMed
-
- Bayless, A. M. , Zapotocny, R. W. , Grunwald, D. J. , Amundson, K. K. , Diers, B. W. , & Bent, A. F. (2018). An atypical N‐ethylmaleimide sensitive factor enables the viability of nematode‐resistant Rhg1 soybeans. Proceedings of the National Academy of Sciences of the United States of America, 115, E4512–E4521. - PMC - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
