

How structural subtleties lead to molecular diversity for the type III polyketide synthases
- PMID: 31471316
- PMCID: PMC6791334
- DOI: 10.1074/jbc.REV119.006129
How structural subtleties lead to molecular diversity for the type III polyketide synthases
Abstract
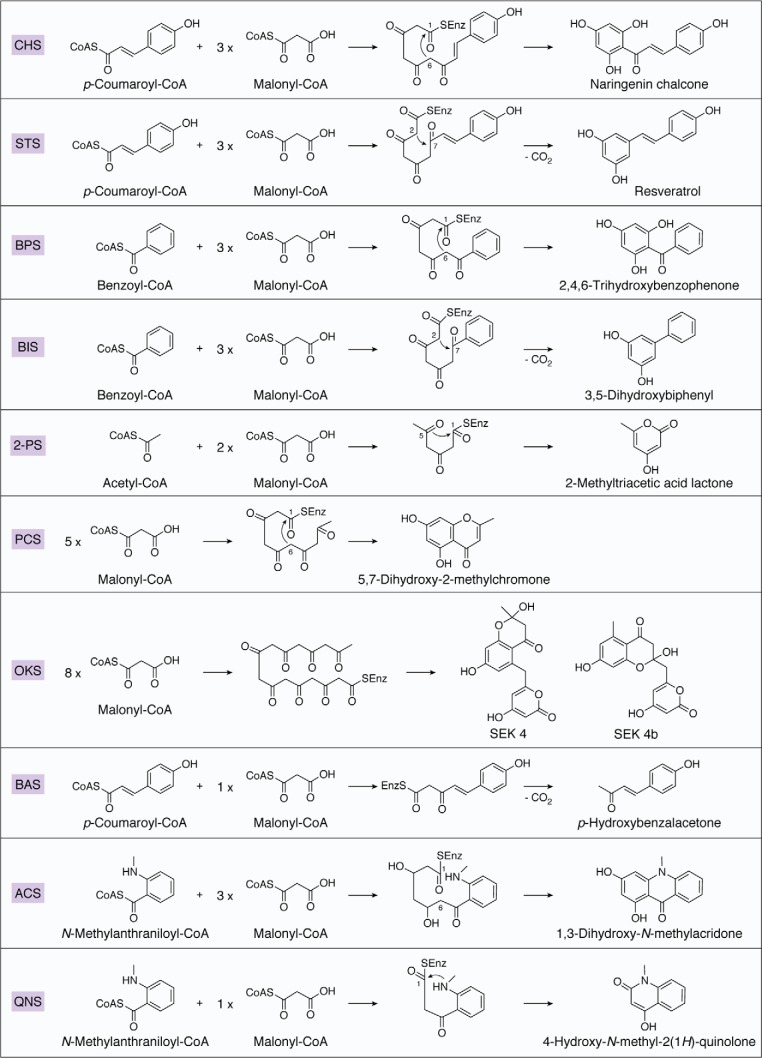
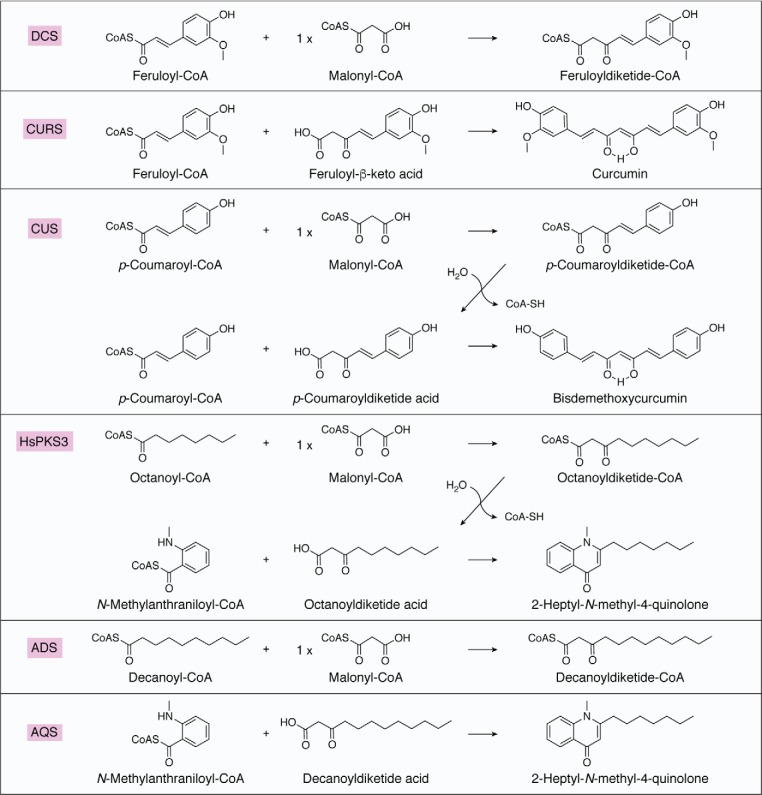
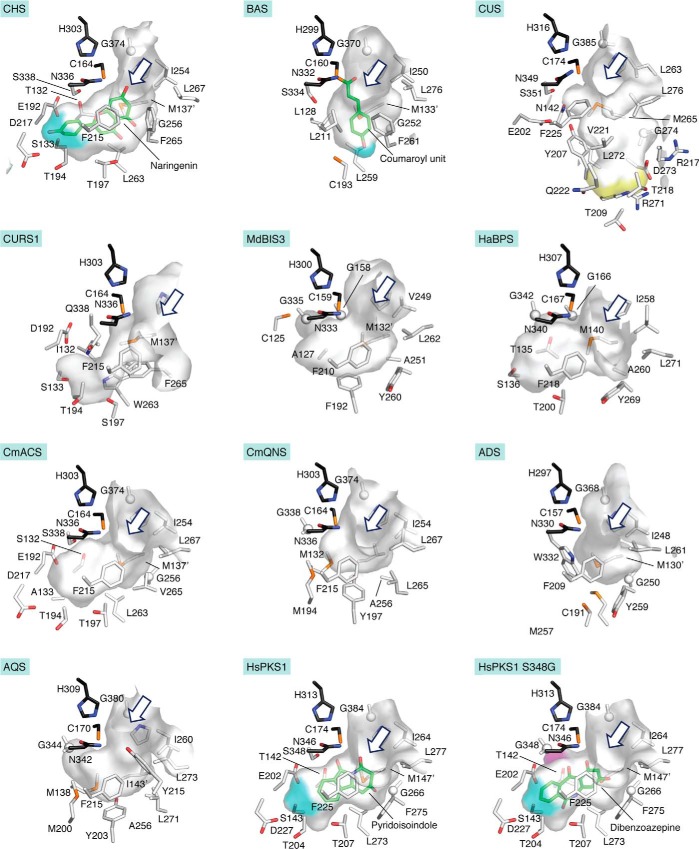
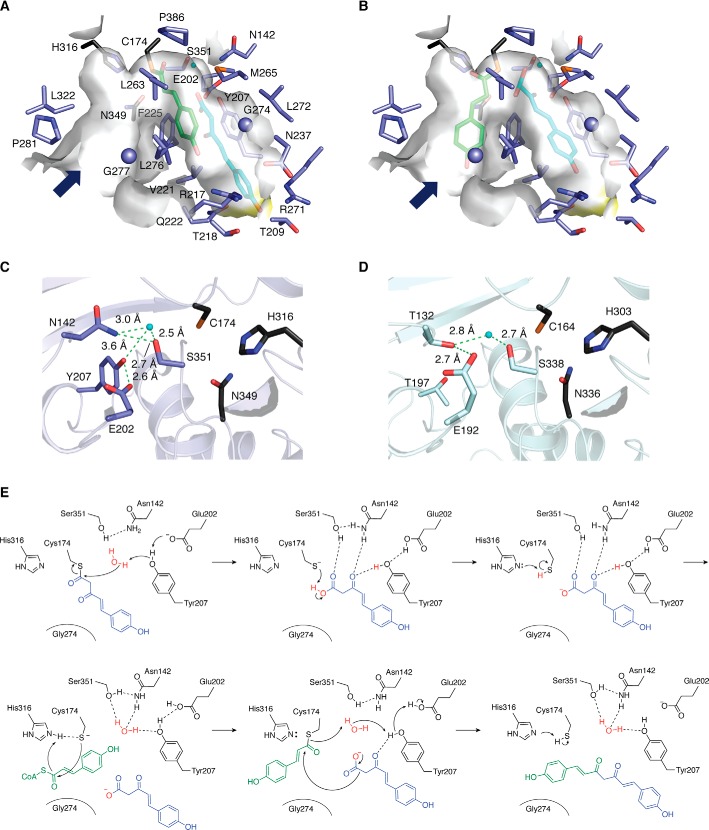
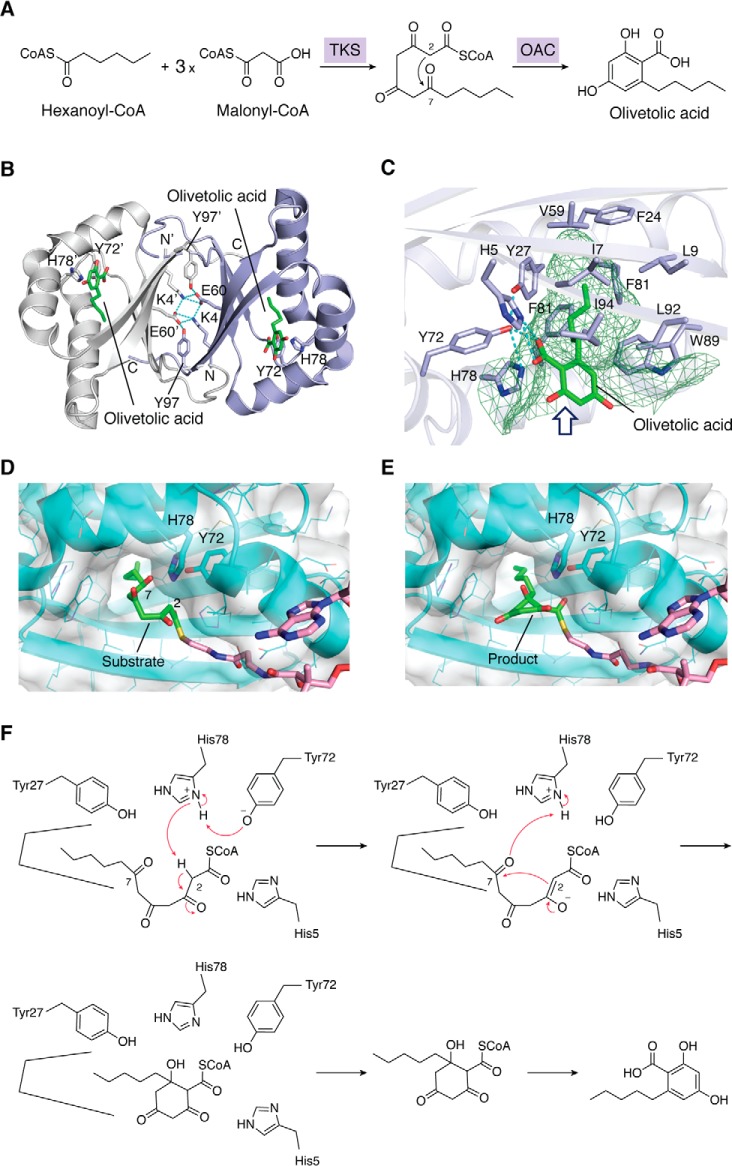
Type III polyketide synthases (PKSs) produce an incredibly diverse group of plant specialized metabolites with medical importance despite their structural simplicity compared with the modular type I and II PKS systems. The type III PKSs use homodimeric proteins to construct the molecular scaffolds of plant polyketides by iterative condensations of starter and extender CoA thioesters. Ever since the structure of chalcone synthase (CHS) was disclosed in 1999, crystallographic and mutational studies of the type III PKSs have explored the intimate structural features of these enzyme reactions, revealing that seemingly minor alterations in the active site can drastically change the catalytic functions and product profiles. New structures described in this review further build on this knowledge, elucidating the detailed catalytic mechanism of enzymes that make curcuminoids, use extender substrates without the canonical CoA activator, and use noncanonical starter substrates, among others. These insights have been critical in identifying structural features that can serve as a platform for enzyme engineering via structure-guided and precursor-directed engineered biosynthesis of plant polyketides. In addition, we describe the unique properties of the recently discovered "second-generation" type III PKSs that catalyzes the one-pot formation of complex molecular scaffolds from three distinct CoA thioesters or from "CoA-free" substrates, which are also providing exciting new opportunities for synthetic biology approaches. Finally, we consider post-type III PKS tailoring enzymes, which can also serve as useful tools for combinatorial biosynthesis of further unnatural novel molecules. Recent progress in the field has led to an exciting time of understanding and manipulating these fascinating enzymes.
Keywords: biosynthesis; enzyme; enzyme mechanism; enzyme structure; natural product biosynthesis.
© 2019 Morita et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures

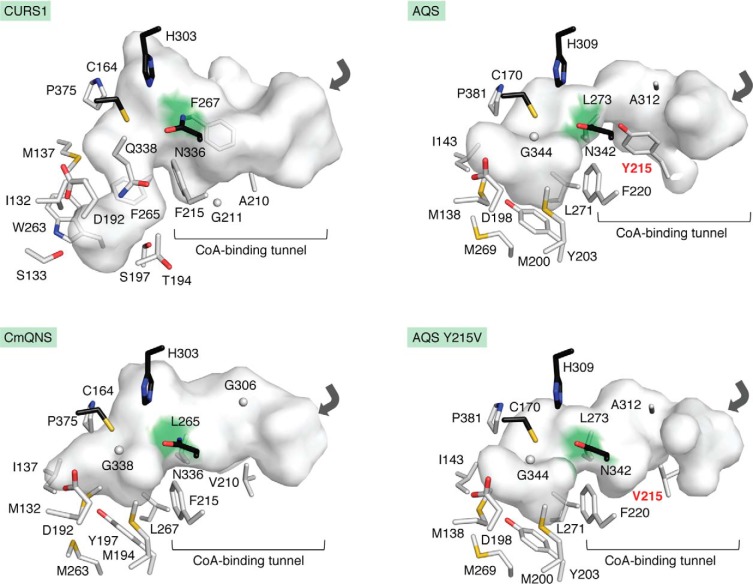
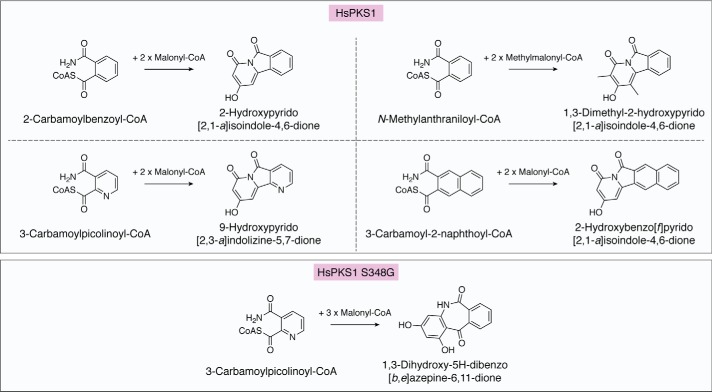
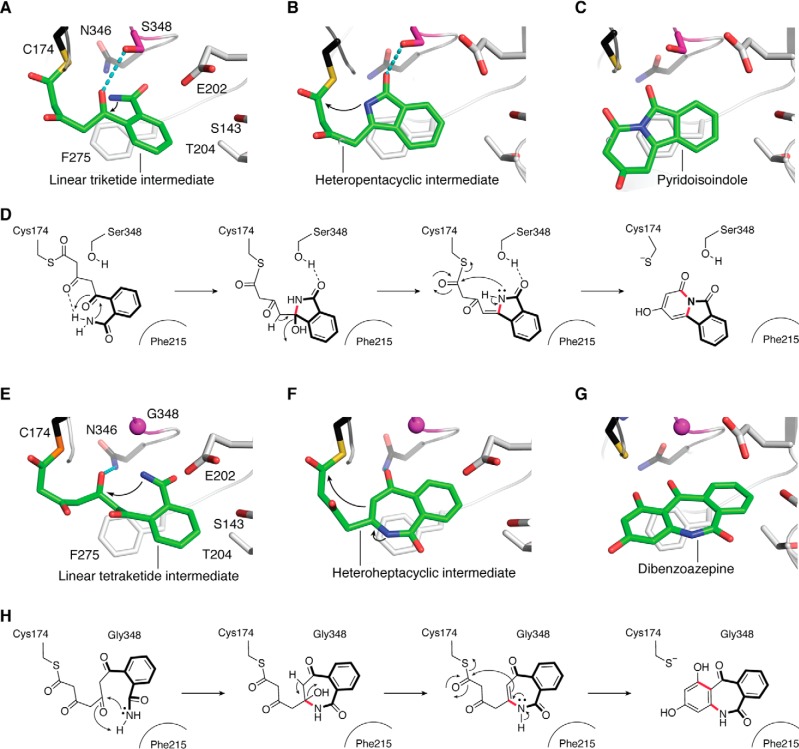
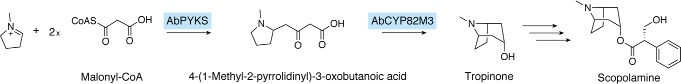
References
-
- Schröder J. (1999) The chalcone-stilbene synthase-type family of condensing enzymes, in Comprehensive Natural Products Chemistry (Sankawa U., ed) pp. 749–771, Elsevier, Oxford, UK
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
