

Complex Oscillatory Waves Emerging from Cortical Organoids Model Early Human Brain Network Development
- PMID: 31474560
- PMCID: PMC6778040
- DOI: 10.1016/j.stem.2019.08.002
Complex Oscillatory Waves Emerging from Cortical Organoids Model Early Human Brain Network Development
Abstract
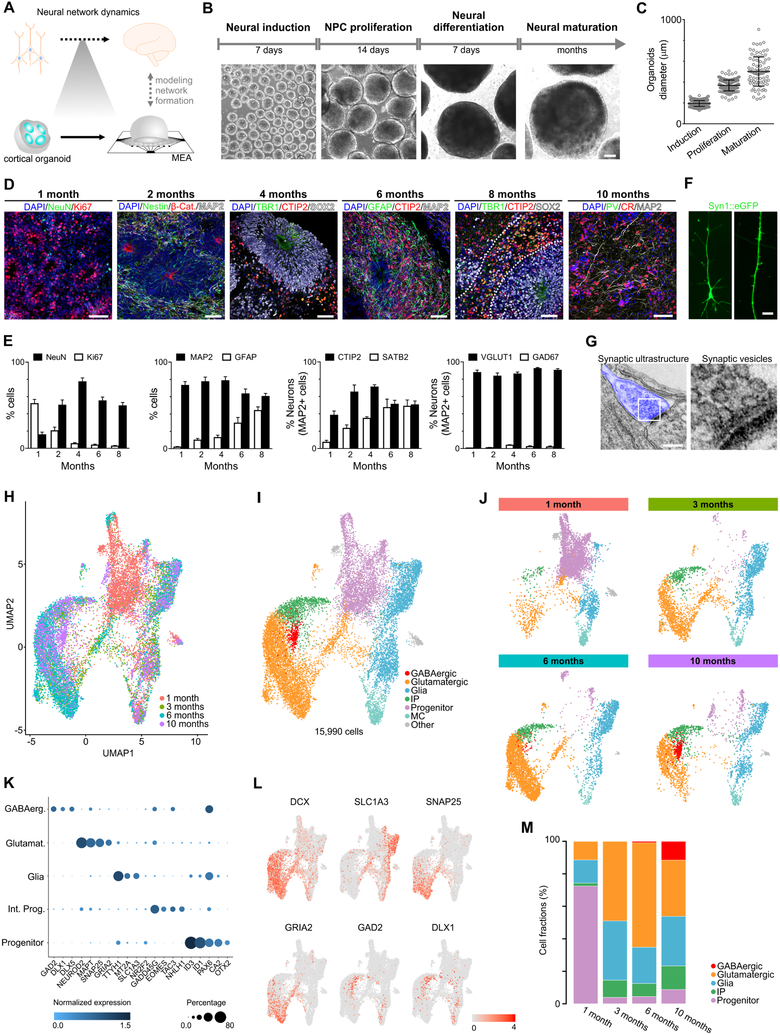
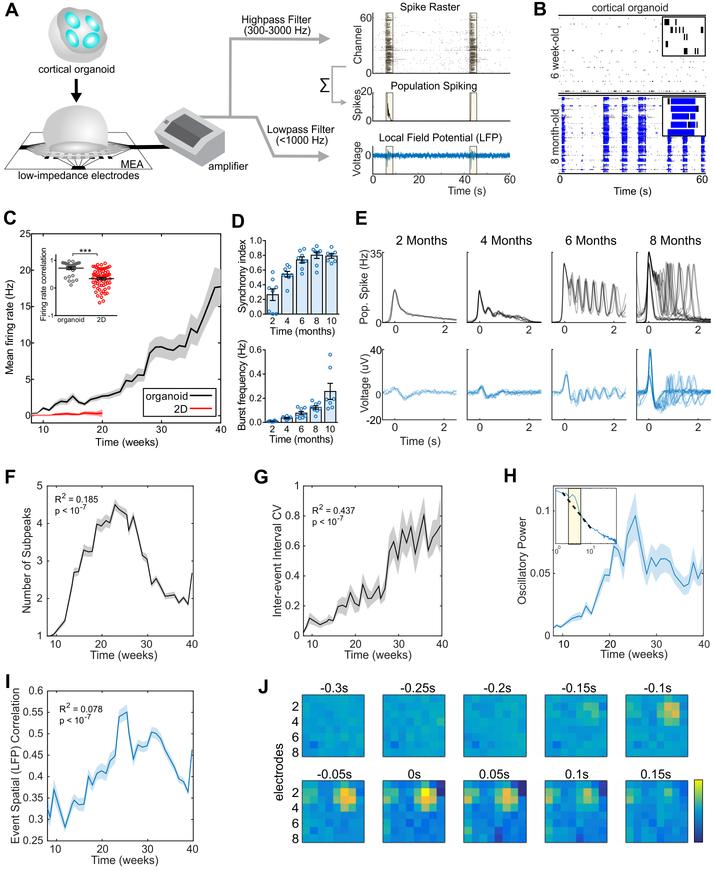
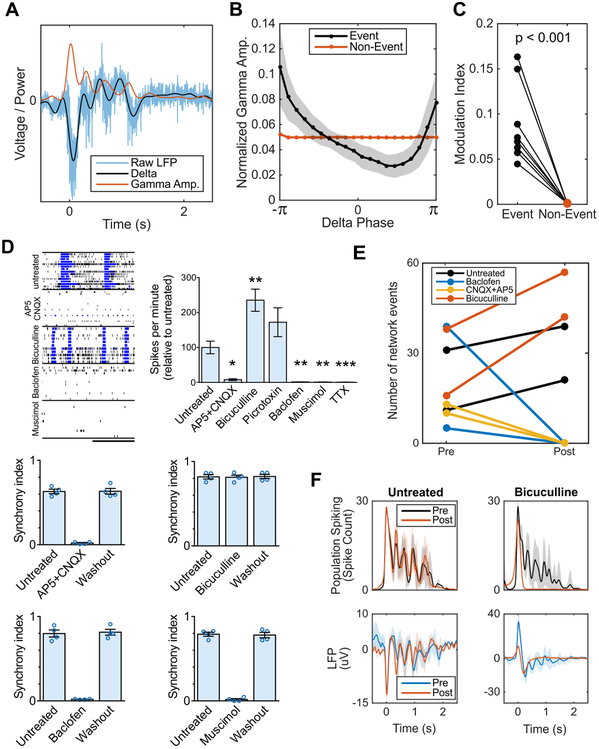
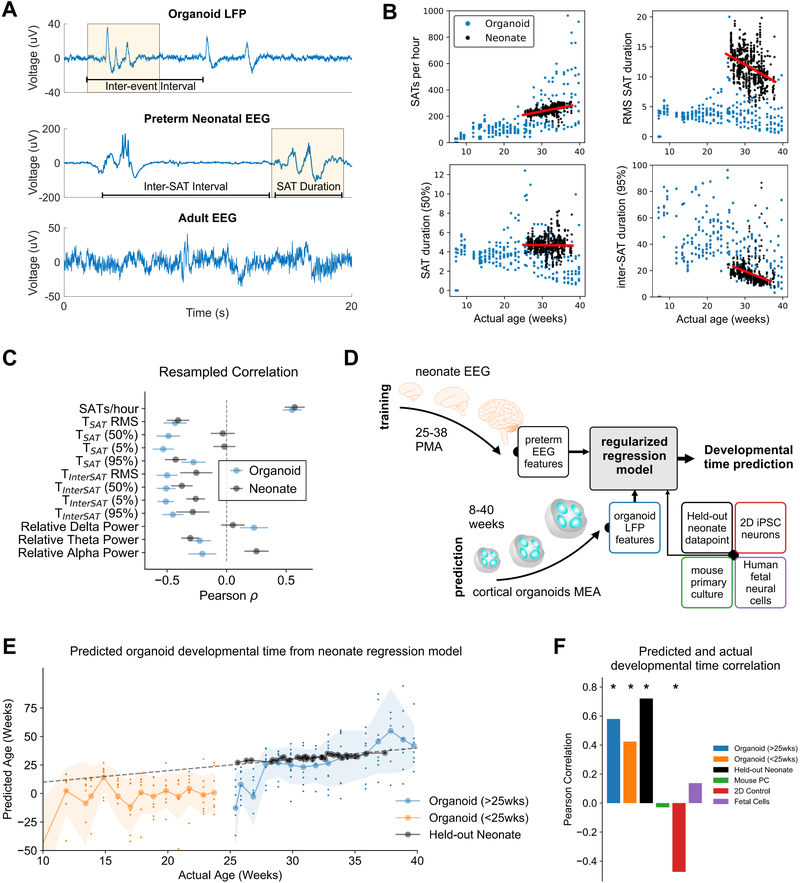
Structural and transcriptional changes during early brain maturation follow fixed developmental programs defined by genetics. However, whether this is true for functional network activity remains unknown, primarily due to experimental inaccessibility of the initial stages of the living human brain. Here, we developed human cortical organoids that dynamically change cellular populations during maturation and exhibited consistent increases in electrical activity over the span of several months. The spontaneous network formation displayed periodic and regular oscillatory events that were dependent on glutamatergic and GABAergic signaling. The oscillatory activity transitioned to more spatiotemporally irregular patterns, and synchronous network events resembled features similar to those observed in preterm human electroencephalography. These results show that the development of structured network activity in a human neocortex model may follow stable genetic programming. Our approach provides opportunities for investigating and manipulating the role of network activity in the developing human cortex.
Keywords: cortical organoids; network oscillations; phase-amplitude coupling; preterm electroencephalography; single-cell transcriptomics; stem cells.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
DECLARATION OF INTERESTS
Dr. Muotri is a co-founder and has equity interest in TISMOO, a company dedicated to genetic analysis focusing on therapeutic applications customized for autism spectrum disorder and other neurological disorders with genetic origins. The terms of this arrangement have been reviewed and approved by the University of California San Diego in accordance with its conflict of interest policies.
Figures
Comment in
-
The Beauty and the Dish: Brain Organoids Go Active.Epilepsy Curr. 2020 Feb 17;20(2):105-107. doi: 10.1177/1535759720901502. eCollection 2020 Mar-Apr. Epilepsy Curr. 2020. PMID: 32313507 Free PMC article. No abstract available.
-
Can lab-grown brains become conscious?Nature. 2020 Oct;586(7831):658-661. doi: 10.1038/d41586-020-02986-y. Nature. 2020. PMID: 33110258 No abstract available.
References
-
- Ben-Ari Y (2001). Developing networks play a similar melody. Trends in Neurosciences 24, 353–360. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
