An official website of the United States government
The .gov means it’s official.
Federal government websites often end in .gov or .mil. Before
sharing sensitive information, make sure you’re on a federal
government site.
The site is secure.
The https:// ensures that you are connecting to the
official website and that any information you provide is encrypted
and transmitted securely.
The development of therapeutic interventions for hearing loss requires fundamental knowledge about the signaling pathways controlling tissue development as well as the establishment of human cell-based assays to validate therapeutic strategies ex vivo Recent advances in the field of stem cell biology and organoid culture systems allow the expansion and differentiation of tissue-specific progenitors and pluripotent stem cells in vitro into functional hair cells and otic-like neurons. We discuss how inner ear organoids have been developed and how they offer for the first time the opportunity to validate drug-based therapies, gene-targeting approaches and cell replacement strategies.
Competing interestsThe authors declare no competing or financial interests.
Figures
Fig. 1.
Schematic of inner ear development. …
Fig. 1.
Schematic of inner ear development. (A) Schematic of embryo development and corresponding tissue…
Fig. 1.
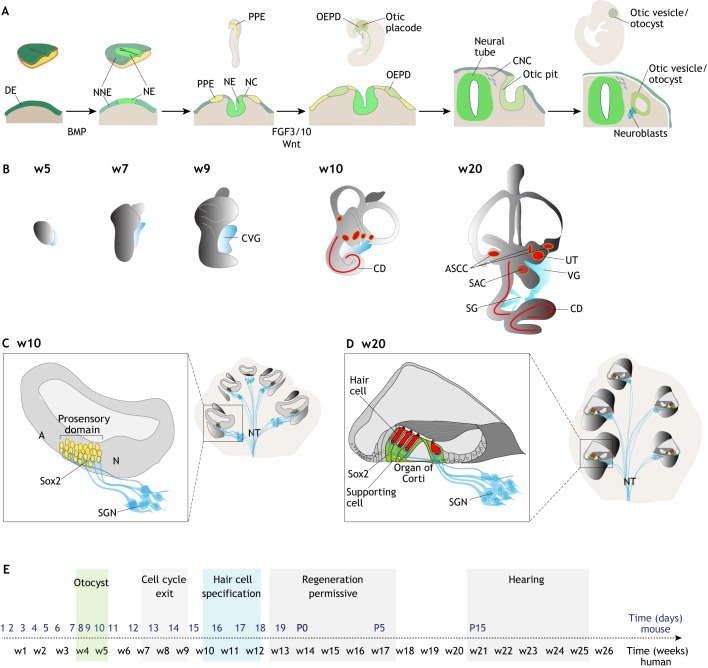
Schematic of inner ear development. (A) Schematic of embryo development and corresponding tissue section, in the cranial portion, to illustrate otic development. Of the three embryonic germ layers, definitive ectoderm (DE) commits to neural fate, giving rise to neural ectoderm (NE). Non-neural ectoderm (NNE) is specified by a lateral-to-medial gradient of BMP signaling. Transient exposure to BMP signaling induces pre-placodal ectoderm (PPE) fate. All cranial placodes, including the otic epibranchial placode domain (OEPD), originate from the PPE. FGF and Wnt promote otic fate. The otic placode invaginates from the surface ectoderm to form the otic pit first, and then the otic vesicle or otocyst. Neuronal progenitors/neuroblasts (blue) delaminate from the otocyst and form the cochlear vestibular ganglion. NC, neural crest; CNC, cranial neural crest. (B) From week 4-5 of human fetal development (E9.5-10.5 mouse) the otocyst grows and gives rise to the components of the inner ear. Epithelial sensory patches are shown in red: three sensory cristae in the ampullae of the semicircular canals (ASCC), two sensory patches in the utricle (UT) and saccule (SAC), and the sensory epithelium in the cochlear duct (CD) contains mechanosensory hair cells. The developing cochlear vestibular ganglion (CVG) is depicted in blue. The vestibular ganglion (VG) neurons innervate the vestibular maculae and cristae. Spiral ganglion (SG) neurons innervate the CD. (C) Schematic of cochlear cross-sections at w10/E14 of development (left) and after maturation (postnatal day 15/w20) (right). The developing prosensory domain in the cochlea is marked by Sox2-positive cells (yellow). Spiral ganglion neurons (SGN) innervate the prosensory domain before hair cell maturation. A, abneural side; N, neural side; NT, nerve trunk. (D) The cochlear prosensory domains differentiate into the organ of Corti. Sensory hair cells are indicated in red, supporting cells in green. (E) Developmental timeline highlighting the steps associated with otocyst formation, cell cycle exit of the cochlear prosensory domain, specification, maturation and functionality of hair cells in the cochlear duct. Human timeline indicated in weeks (w) in black, mouse timeline in days in blue. Postnatal days (P) 0, 5 and 15 are indicated.
Fig. 2.
Preparation of cochlear organoids from…
Fig. 2.
Preparation of cochlear organoids from murine and human tissue-specific progenitors. (A) Organoid derivation…
Fig. 2.
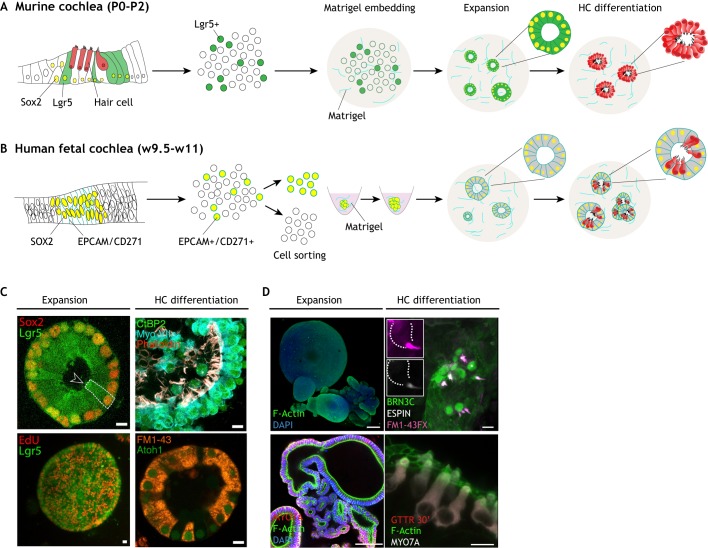
Preparation of cochlear organoids from murine and human tissue-specific progenitors. (A) Organoid derivation from murine Lgr5-positive cochlear supporting cells, isolated from early postnatal mice (P0-P2) using an Lgr5-EGFP reporter line. The sensory epithelium is dissociated to a single cell suspension and embedded in Matrigel. A first phase of expansion of Lgr5-positive progenitors is followed by differentiation, induced by Wnt activation and Notch inhibition, resulting in conversion of Lgr5-positive supporting cells (green) to hair cells (HC; red). (B) Organoid derivation from human fetal prosensory domain cells. The whole cochlea is mechanically and enzymatically dissociated into a single cell suspension. EPCAM and CD271 (green) are used as surface markers to isolate Sox2-positive cells from the prosensory domain. Cells are sorted by flow cytometry and aggregated in round-bottom 96-well plates, then embedded in Matrigel and expanded for 2-3 weeks. Differentiation is induced in co-culture with the mesenchymal/neuronal progenitor pool and growth factor withdrawal for an additional 2-3 weeks. This results in organoids that contain supporting cells (gray) and hair cells (red). (C) Representative examples of cochlear organoids as in McLean et al. (2017). Immunostaining of the organoids during the expansion and differentiation phases. During expansion (left), epithelial cells in the cysts co-express Sox2 and Lgr5 (GFP) (upper panel) and are highly proliferative (lower panel). Upon differentiation (right), they express hair cell markers such as Myo7a, CtBP2 and F-actin-positive bundles (upper panel). Cells also express Atoh1 and are capable of taking up the FM1-43 dye, suggesting active MET channels (lower panel). The dashed line indicates the border of a single supporting cell. The arrowhead points at the apical surface, facing the lumen. (D) Representative examples of cochlear organoids from human fetal tissue as in Roccio et al. (2018). Immunostaining of the organoids during the expansion and differentiation phases. During the expansion phase (left), sorted cells grow as epithelial structures expressing different cochlear duct and prosensory domain markers and lack differentiation signs (MYO7A, F-actin bundles). After differentiation (right), patches of hair cells expressing BRN3C, espin and MYO7A are found. In vitro-generated hair cells show functional MET channels as detected by FM1-43 uptake (upper panel) and GTTR uptake (lower panel). The dashed line indicates the contour of a single hair cell. The asterisk marks the cell enlarged in the insets. Scale bars: 15 μm in C; 100 μm in D (expansion); 10 μm in D (differentiation).
Fig. 3.
Pluripotent stem cell - derived…
Fig. 3.
Pluripotent stem cell - derived inner ear organoids. (A). Schematic of in vitro …
Fig. 3.
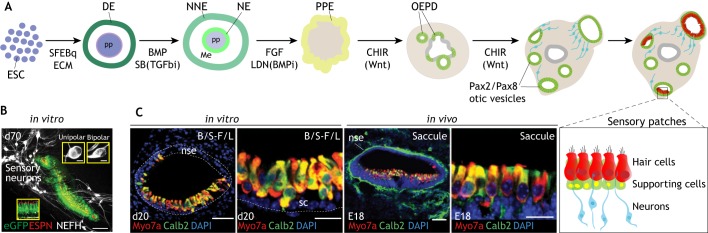
Pluripotent stem cell-derived inner ear organoids. (A). Schematic of in vitro guided organogenesis from pluripotent stem cells to otic tissue adapted from the mouse and human protocols. Embryonic stem cells (ESC) are aggregated with serum-free embryoid body-quick (SFEBq) methods and embedded in Matrigel (ECM) to induce definitive ectoderm (DE) at the surface of the aggregate. Non-neural ectoderm (NNE) is induced by BMP4 addition (or its endogenous production) and concomitant inhibition of mesendoderm fate with a TGFβ inhibitor (SB; SB451542). Pre-placodal ectoderm (PPE) fate is promoted by the addition of FGF (Fgf2) and BMP inhibitor (LDN; LDN193189). Otic epibranchial placode (OEPD) and otic fate are promoted by activation of Wnt signaling with GSK3β inhibitor, CHIR99021 (CHIR). Pax2/Pax8 otic vesicles form within the aggregate and further develop into Jag1+/Sox2+ prosensory patches (not shown) and later into sensory patches containing hair cells (red) and supporting cells (green), as well as otic sensory neurons (blue). (B) Representative images from Koehler et al. (2017) showing inner ear organoids derived from the differentiation of hESCs. The mechanosensitive hair cells are labeled by Atoh-1 GFP (green), hair bundles are stained for espin (red) and co-differentiated sensory neurons are shown in white (NEFH) at day 70 (d70) of in vitro differentiation. (C) Comparison of in vitro-generated sensory hair cells (20 days in vitro; d20) from mESCs and hair cells resident in the murine saccule at E18 of mouse embryonic development. Hair cells are immunostained for Myo7a (red) and calbindin 2 (calb2, green). Figures from Koehler et al. (2013). Dotted lines delineate the border of the vesicle and the non-sensory epithelium (nse). B/S, BMP/SB; F/L, FGF/LDN; Me, mesoderm; NE, neural ectoderm; pp, pluripotent; sc, supporting cells. Scale bars: 100 μm in B (5 μm in inserts); 50 μm in C (1st and 3rd panels); 25 μm in C (2nd and 4th panels).
Abbas L. and Rivolta M. N. (2015). Aminoglycoside ototoxicity and hair cell ablation in the adult gerbil: a simple model to study hair cell loss and regeneration. Hear. Res. 325, 12-26. 10.1016/j.heares.2015.03.002
-
DOI
-
PMC
-
PubMed
Abbas L. and Rivolta M. N. (2019). The use of animal models to study cell transplantation in neuropathic hearing loss. Hear. Res. 377, 72-87. 10.1016/j.heares.2019.03.014
-
DOI
-
PubMed
Alsina B. and Whitfield T. T. (2017). Sculpting the labyrinth: morphogenesis of the developing inner ear. Semin. Cell Dev. Biol. 65, 47-59. 10.1016/j.semcdb.2016.09.015
-
DOI
-
PubMed
Appler J. M. and Goodrich L. V. (2011). Connecting the ear to the brain: molecular mechanisms of auditory circuit assembly. Prog. Neurobiol. 93, 488-508. 10.1016/j.pneurobio.2011.01.004
-
DOI
-
PMC
-
PubMed
Atkinson P. J., Wise A. K., Flynn B. O., Nayagam B. A. and Richardson R. T. (2014). Hair cell regeneration after ATOH1 gene therapy in the cochlea of profoundly deaf adult guinea pigs. PLoS ONE 9, e102077 10.1371/journal.pone.0102077
-
DOI
-
PMC
-
PubMed