

Control of DNA replication timing in the 3D genome
- PMID: 31477886
- PMCID: PMC11567694
- DOI: 10.1038/s41580-019-0162-y
Control of DNA replication timing in the 3D genome
Abstract
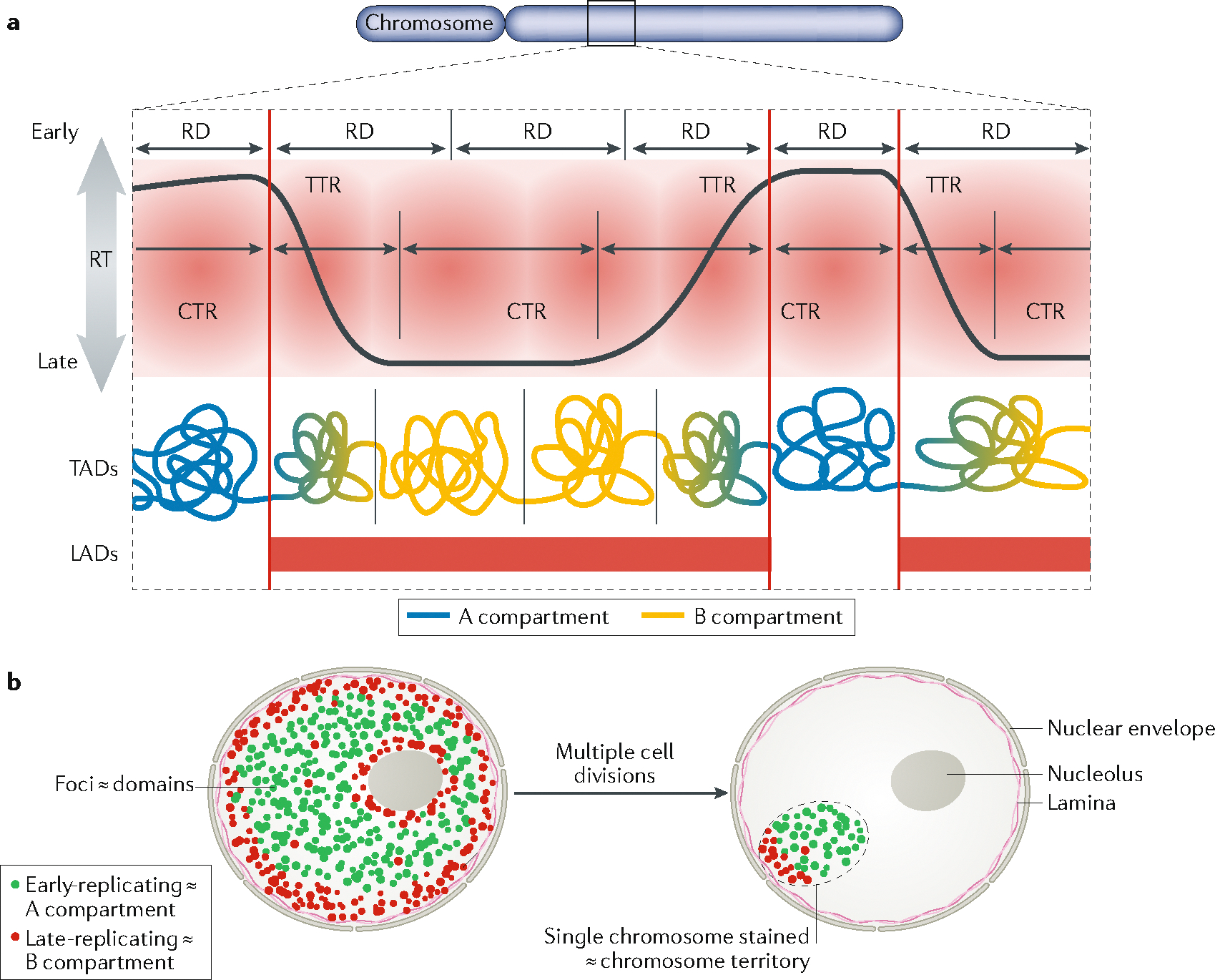
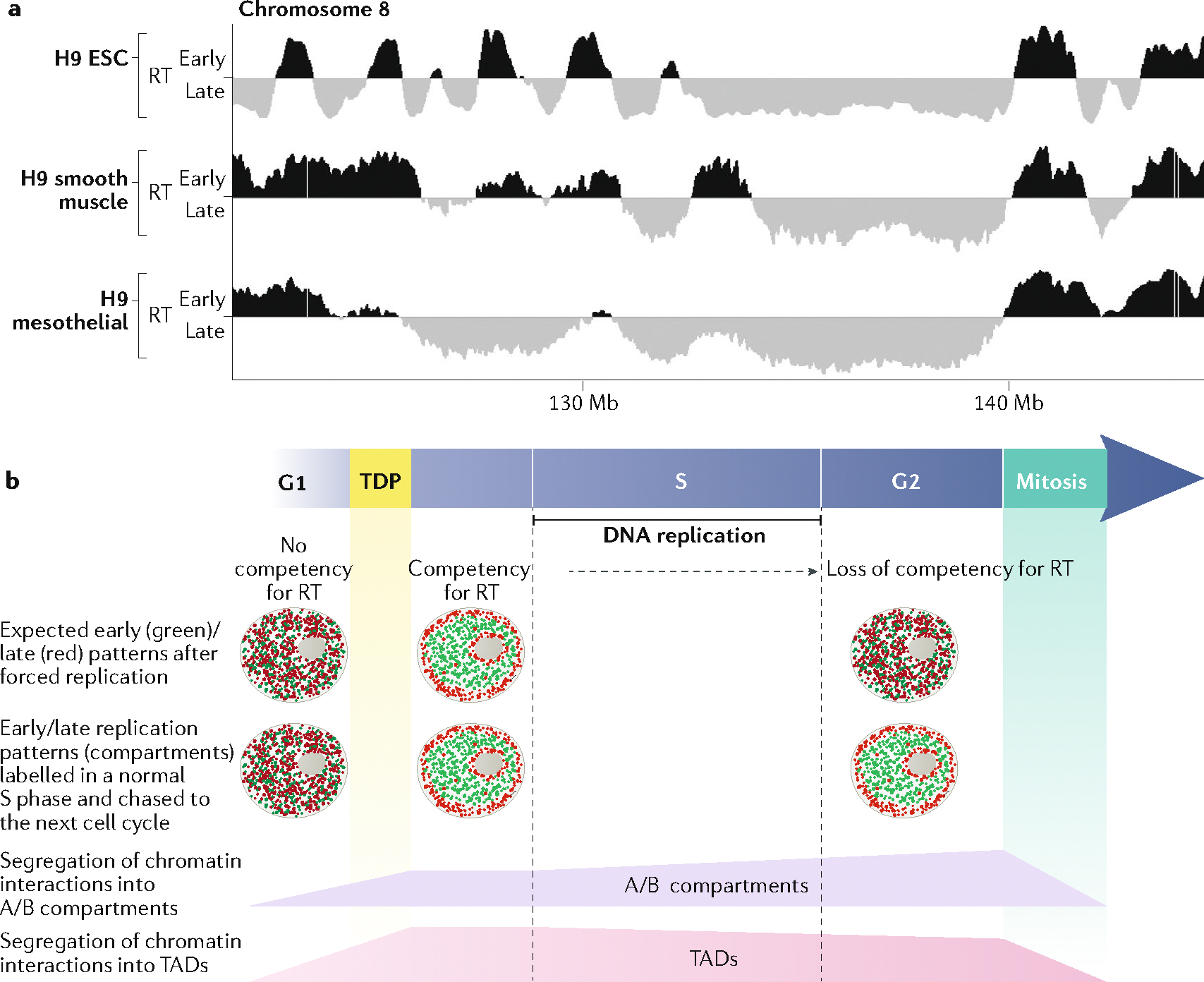
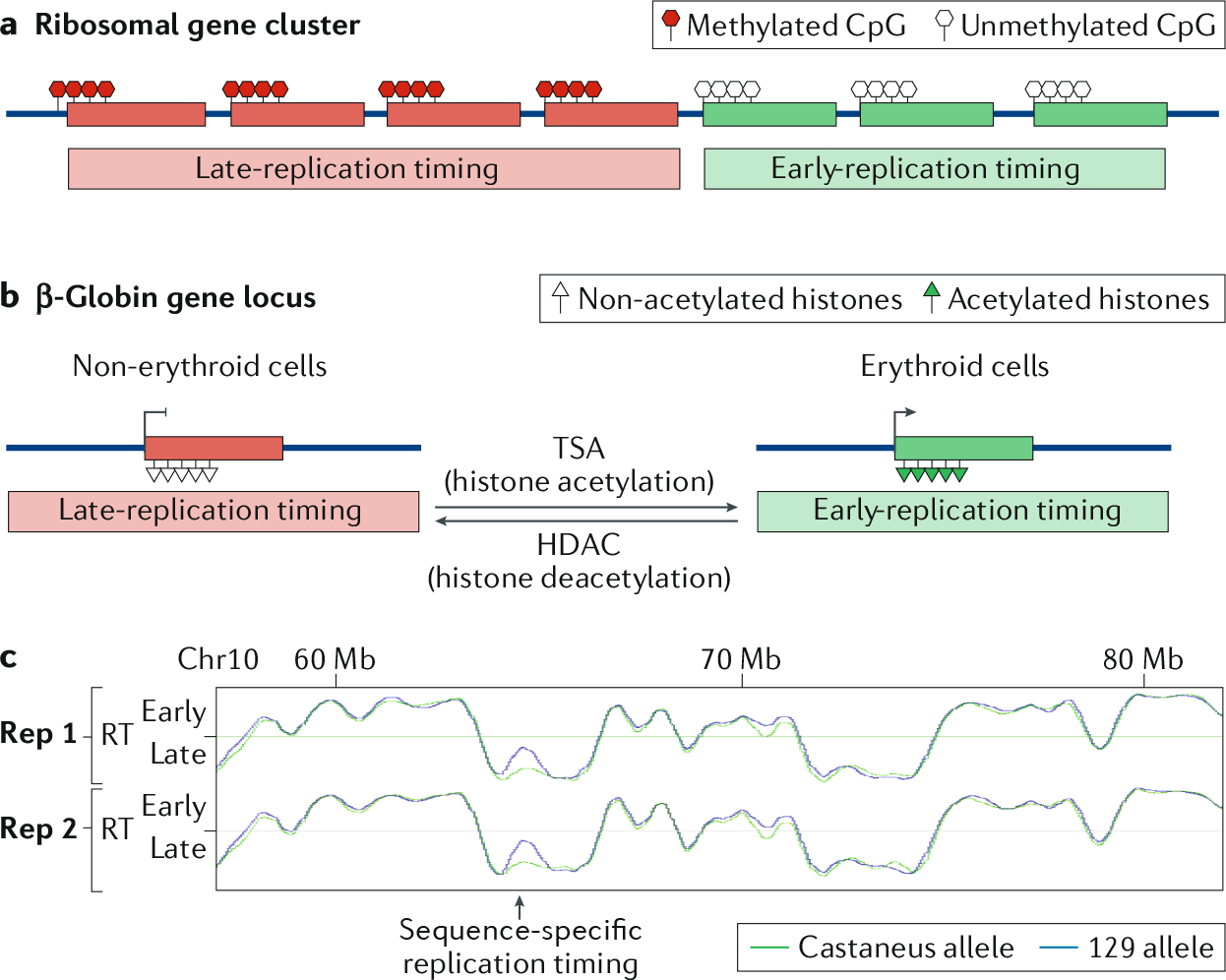
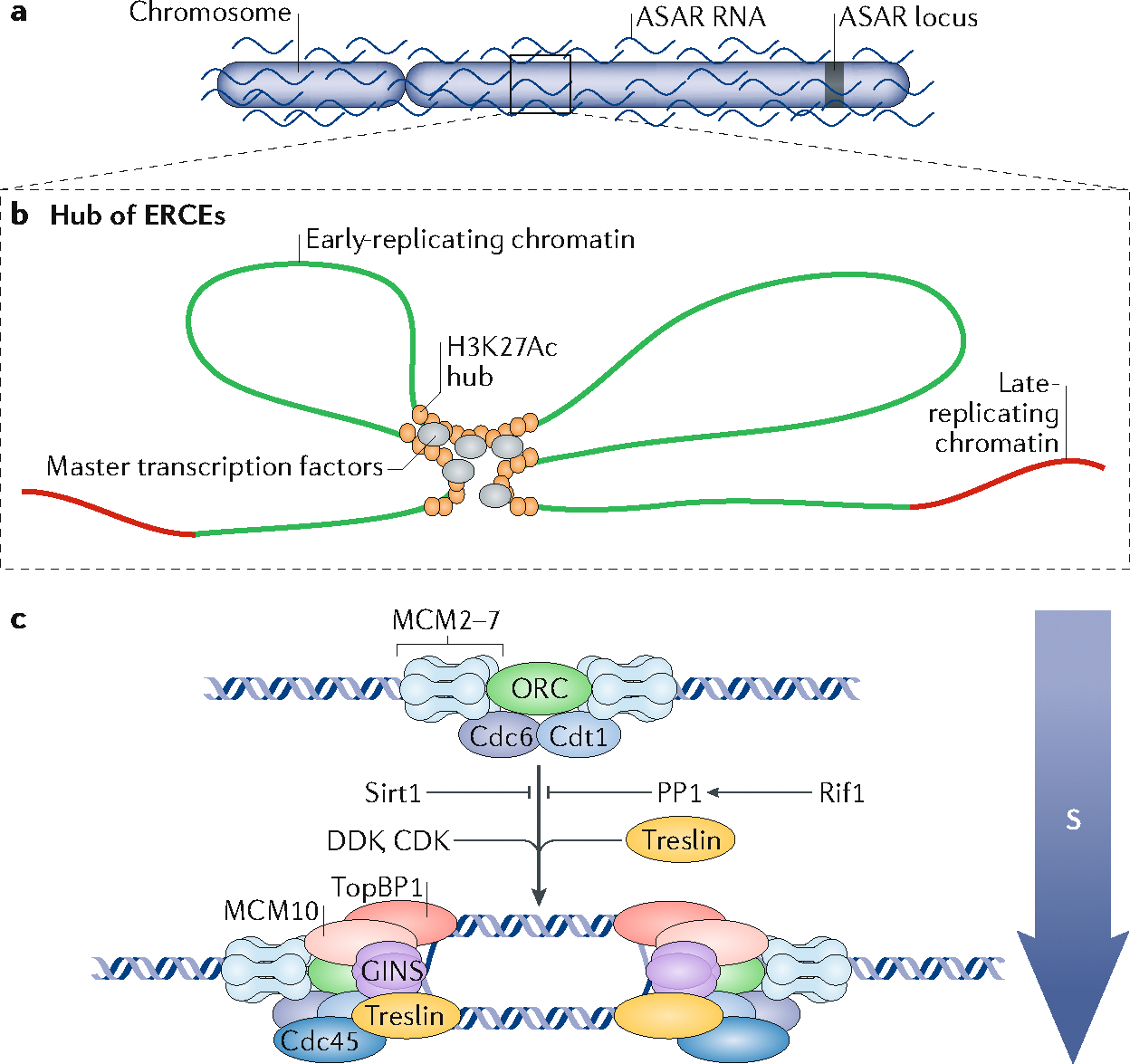
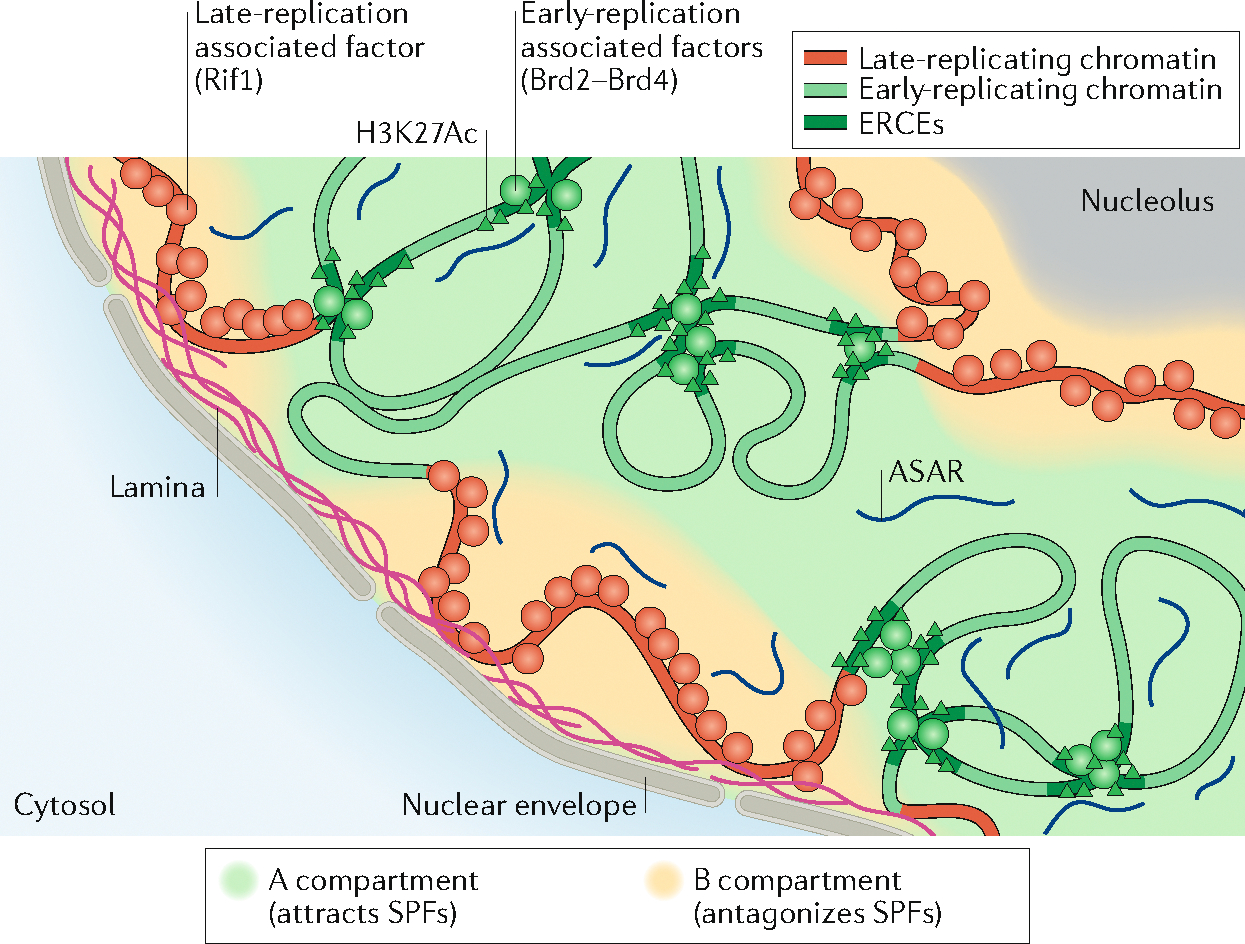
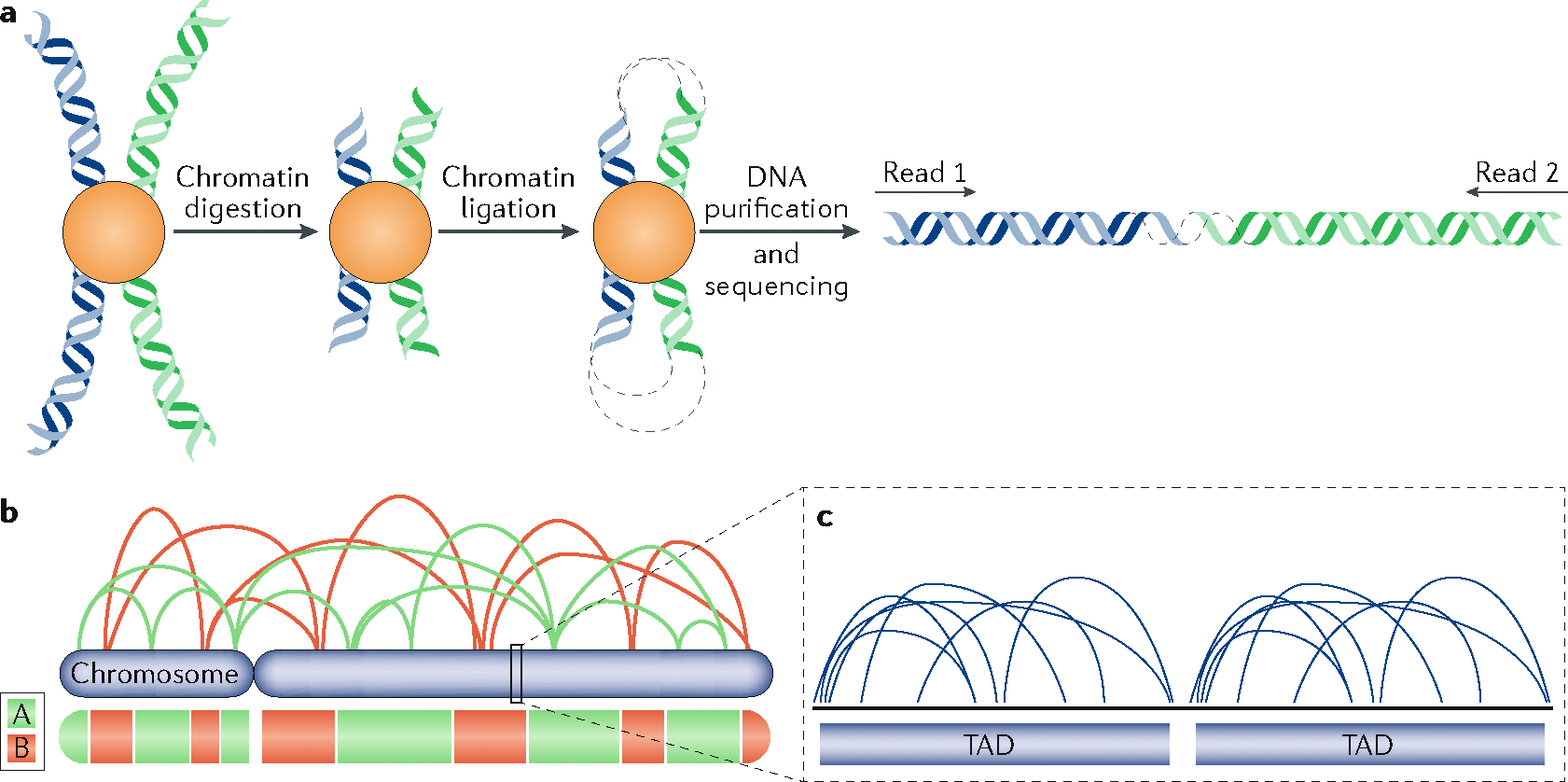
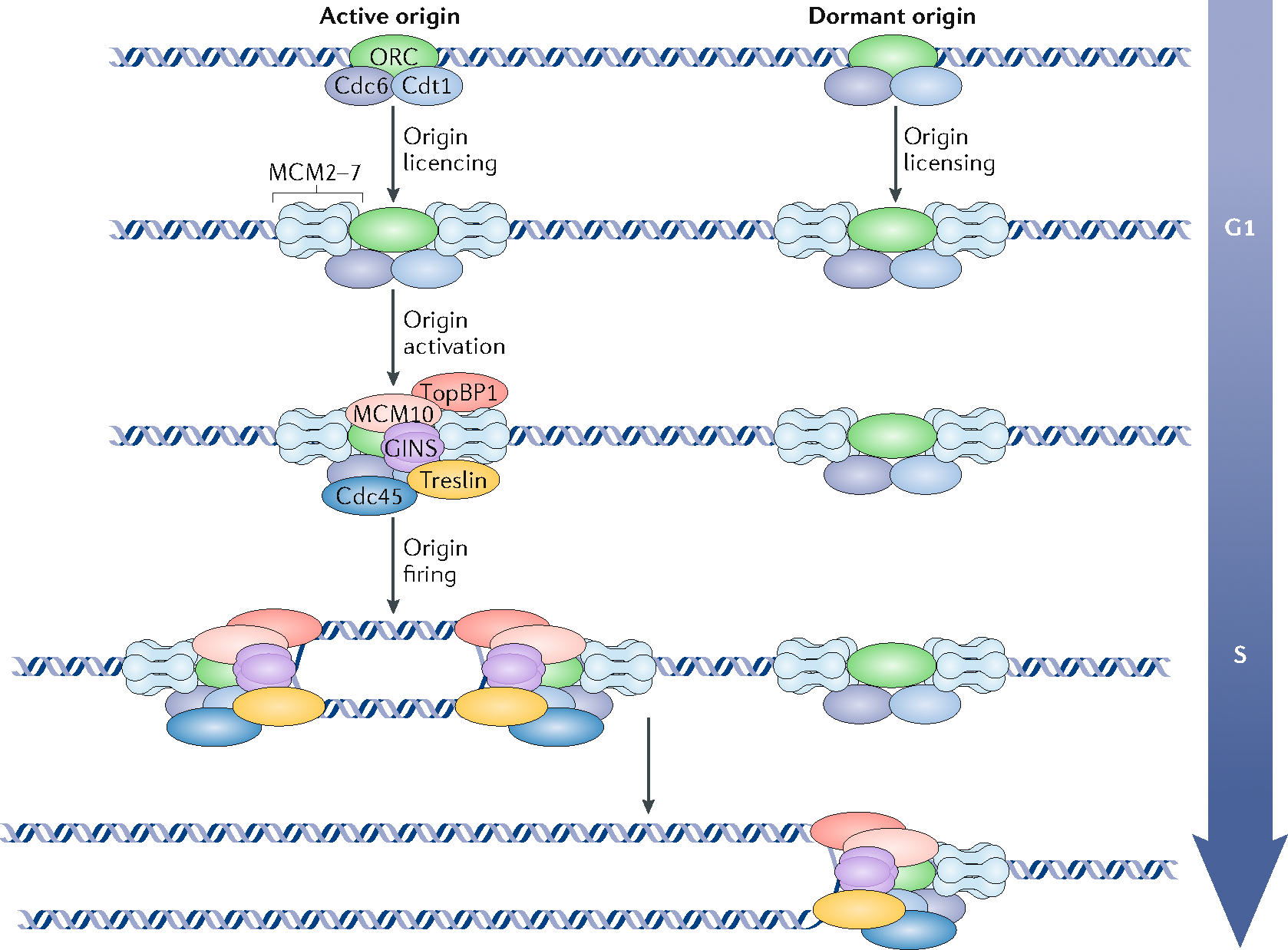
The 3D organization of mammalian chromatin was described more than 30 years ago by visualizing sites of DNA synthesis at different times during the S phase of the cell cycle. These early cytogenetic studies revealed structurally stable chromosome domains organized into subnuclear compartments. Active-gene-rich domains in the nuclear interior replicate early, whereas more condensed chromatin domains that are largely at the nuclear and nucleolar periphery replicate later. During the past decade, this spatiotemporal DNA replication programme has been mapped along the genome and found to correlate with epigenetic marks, transcriptional activity and features of 3D genome architecture such as chromosome compartments and topologically associated domains. But the causal relationship between these features and DNA replication timing and the regulatory mechanisms involved have remained an enigma. The recent identification of cis-acting elements regulating the replication time and 3D architecture of individual replication domains and of long non-coding RNAs that coordinate whole chromosome replication provide insights into such mechanisms.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
