

Chimeric antigen receptor-induced BCL11B suppression propagates NK-like cell development
- PMID: 31479431
- PMCID: PMC6877334
- DOI: 10.1172/JCI126350
Chimeric antigen receptor-induced BCL11B suppression propagates NK-like cell development
Abstract
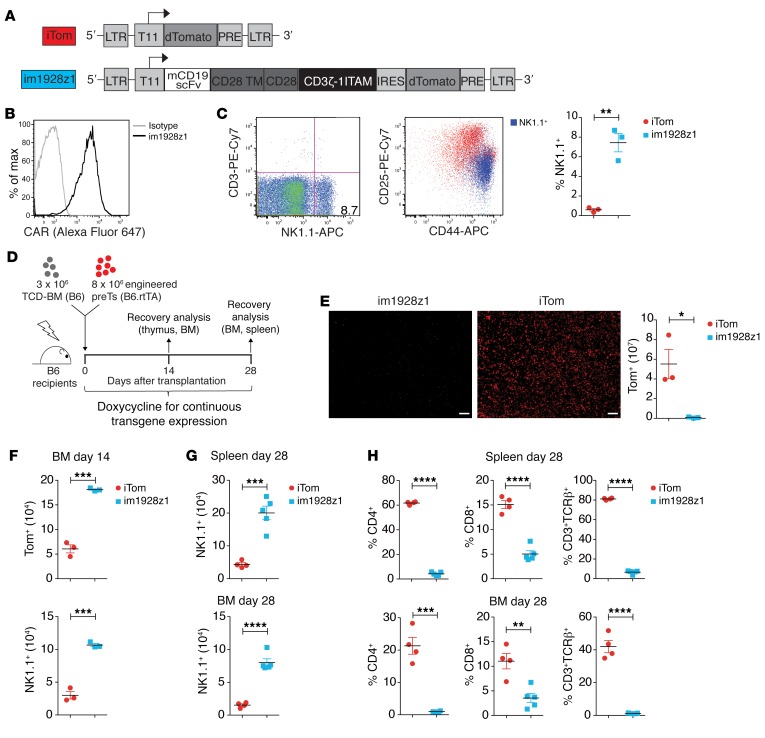
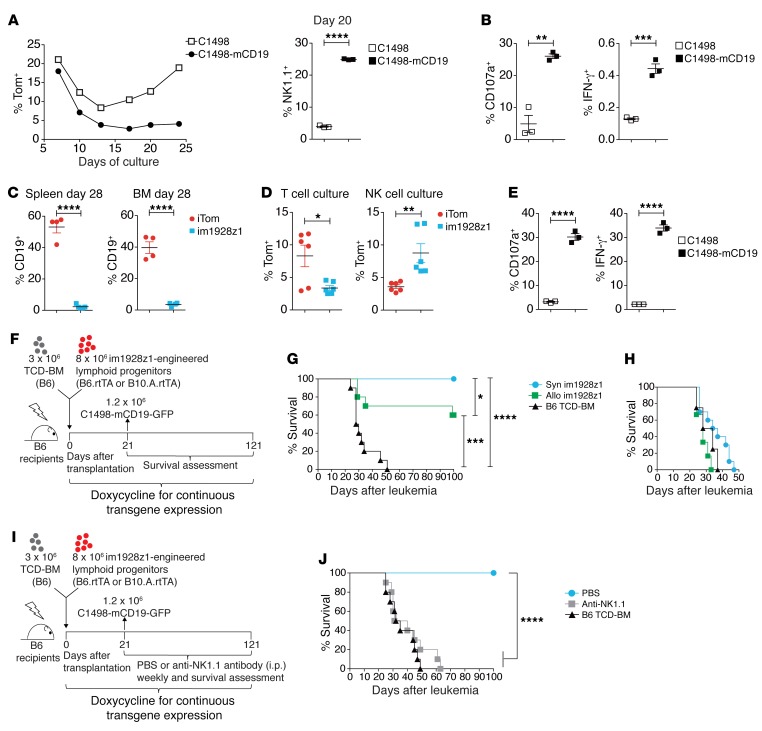
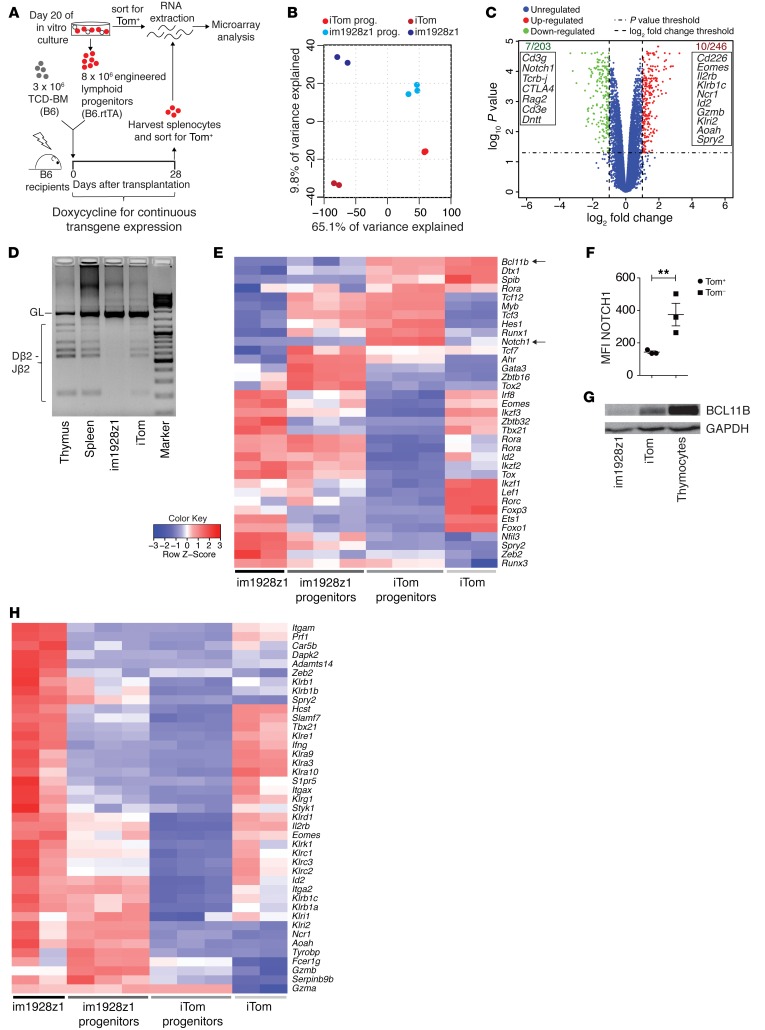
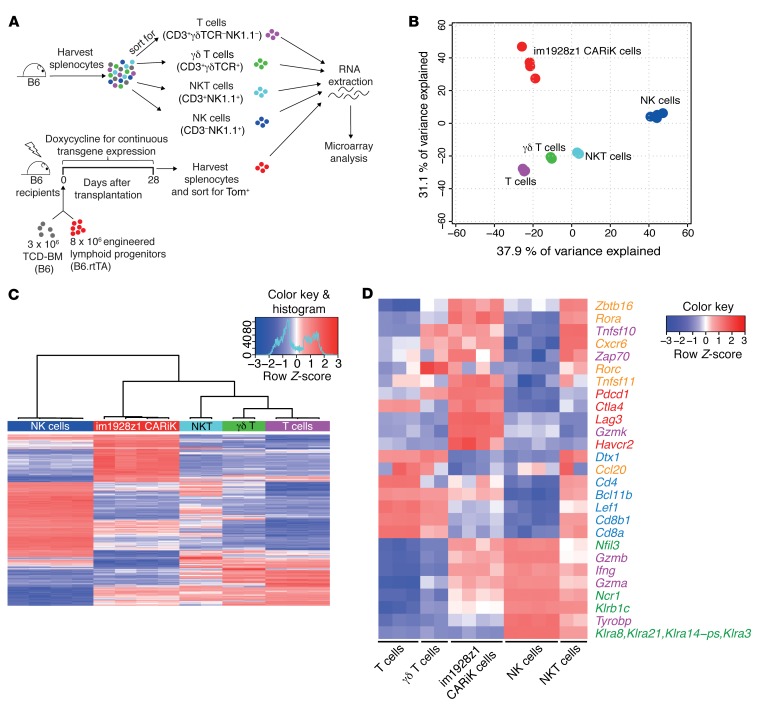
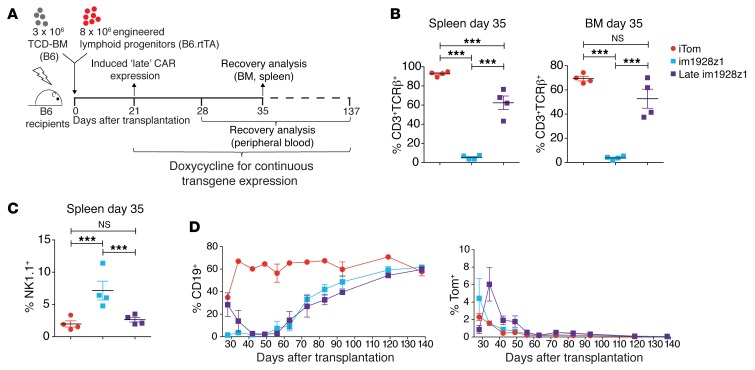
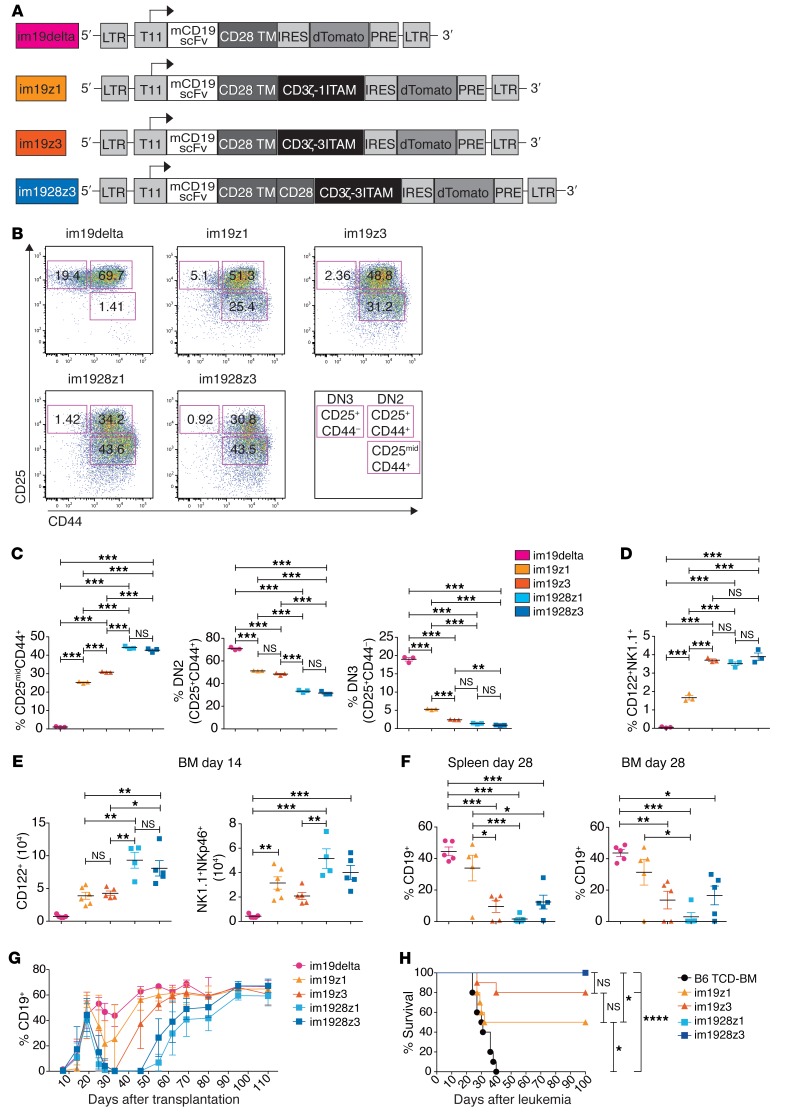
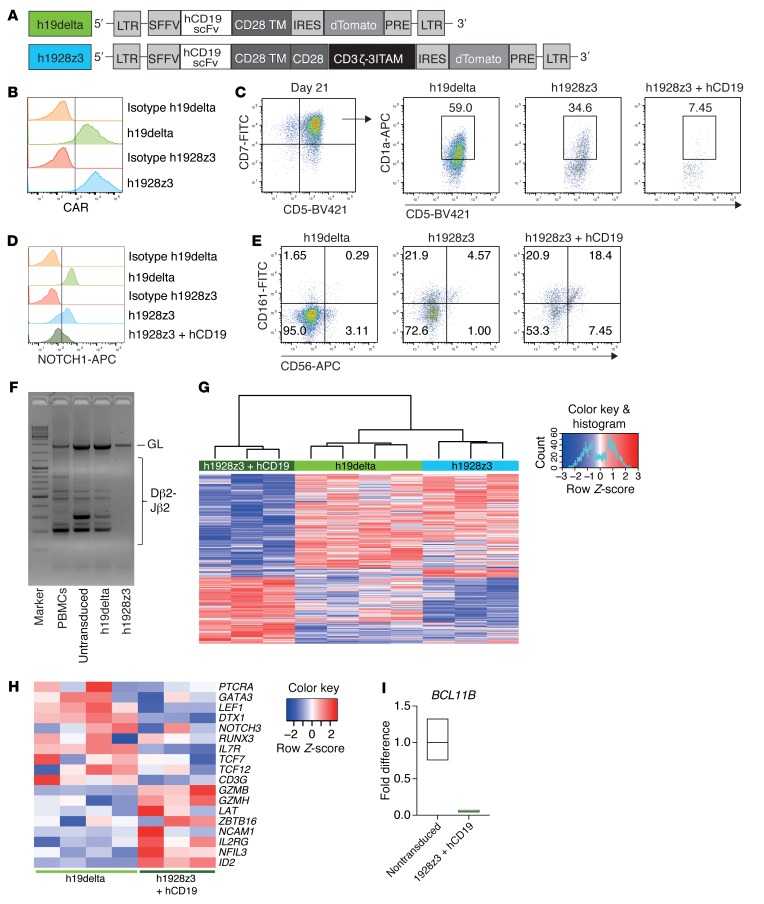
The transcription factor B cell CLL/lymphoma 11B (BCL11B) is indispensable for T lineage development of lymphoid progenitors. Here, we show that chimeric antigen receptor (CAR) expression during early phases of ex vivo generation of lymphoid progenitors suppressed BCL11B, leading to suppression of T cell-associated gene expression and acquisition of NK cell-like properties. Upon adoptive transfer into hematopoietic stem cell transplant recipients, CAR-expressing lymphoid progenitors differentiated into CAR-induced killer (CARiK) cells that mediated potent antigen-directed antileukemic activity even across MHC barriers. CD28 and active immunoreceptor tyrosine-based activation motifs were critical for a functional CARiK phenotype. These results give important insights into differentiation of murine and human lymphoid progenitors driven by synthetic CAR transgene expression and encourage further evaluation of ex vivo-generated CARiK cells for targeted immunotherapy.
Keywords: Immunology; Immunotherapy; Leukemias; T cell development; Transplantation.
Conflict of interest statement
Figures
Comment in
-
Adding chimeric antigen receptor-induced killer cells to the medical oncology shelf.J Clin Invest. 2019 Dec 2;129(12):5077-5078. doi: 10.1172/JCI132536. J Clin Invest. 2019. PMID: 31638599 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
