

Syndecan-Mediated Ligation of ECM Proteins Triggers Proliferative Arrest of Disseminated Tumor Cells
- PMID: 31481497
- PMCID: PMC7685526
- DOI: 10.1158/0008-5472.CAN-19-1165
Syndecan-Mediated Ligation of ECM Proteins Triggers Proliferative Arrest of Disseminated Tumor Cells
Abstract
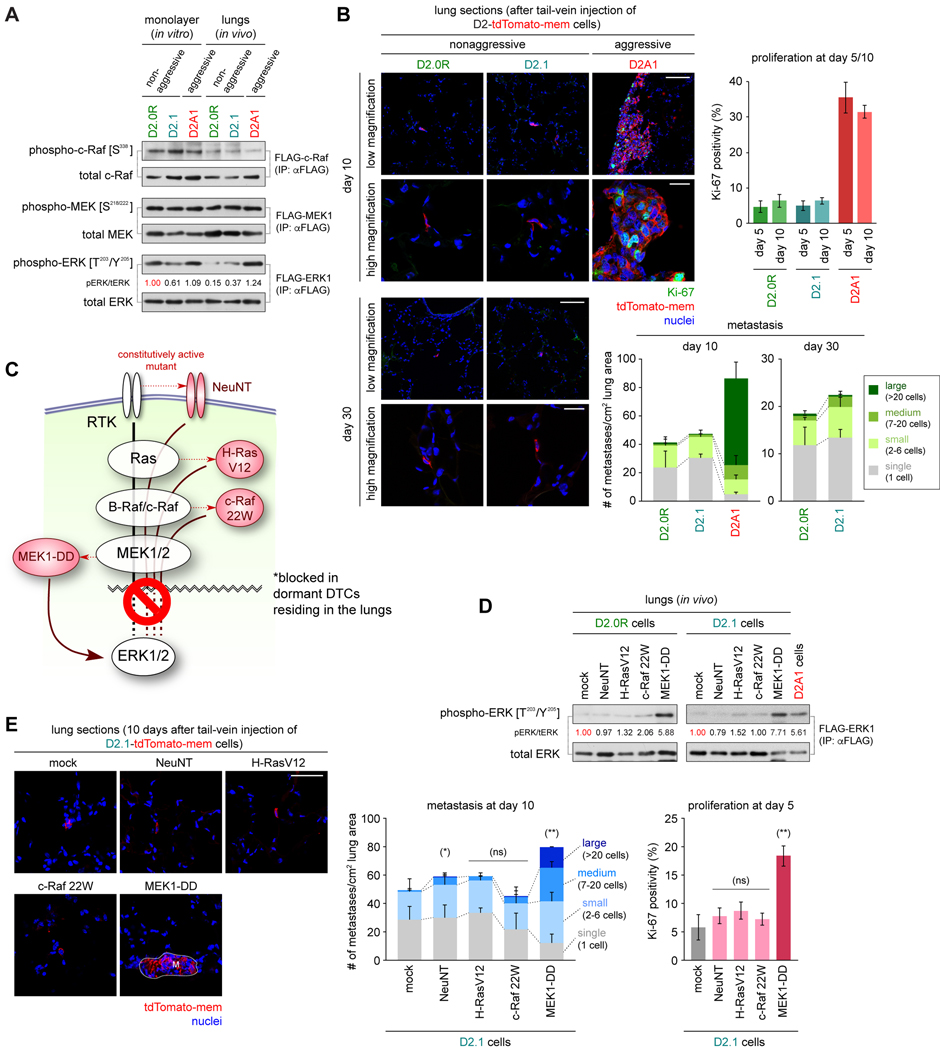
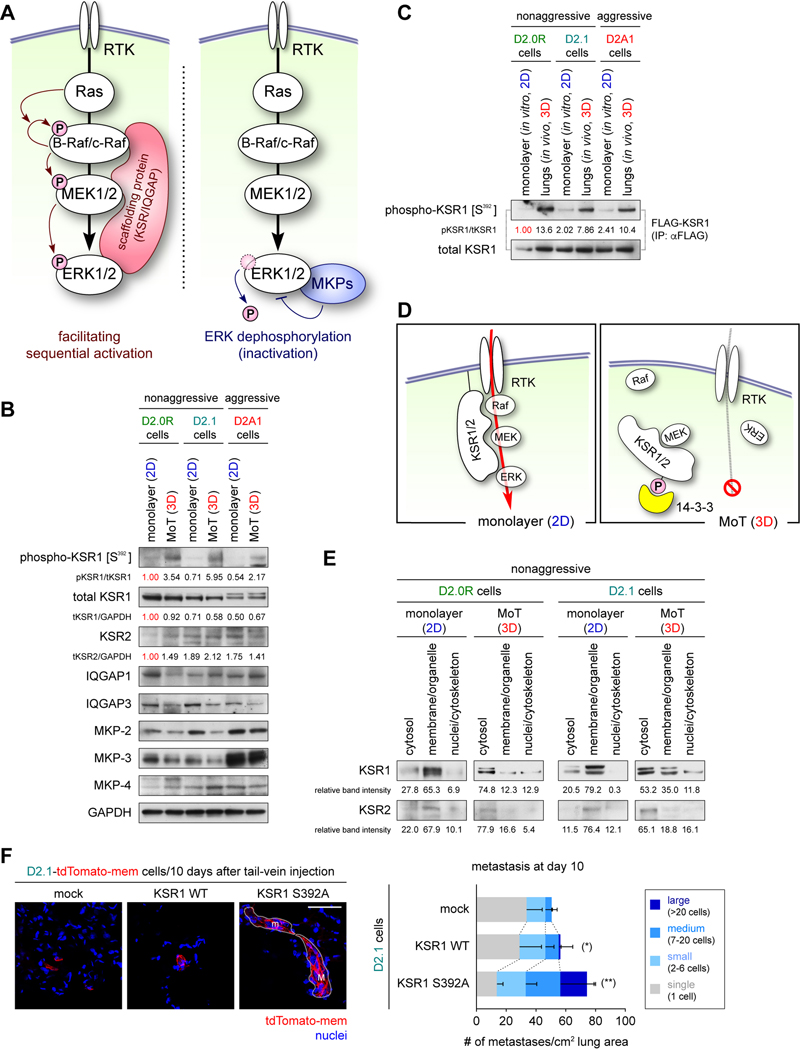
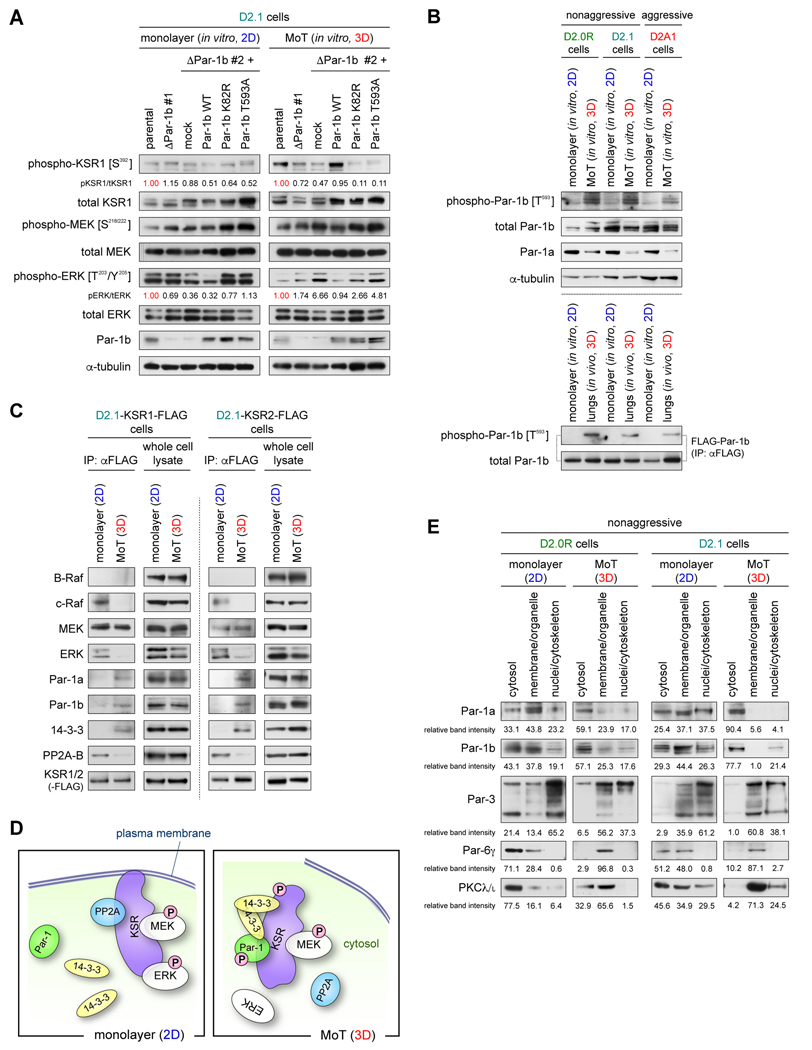
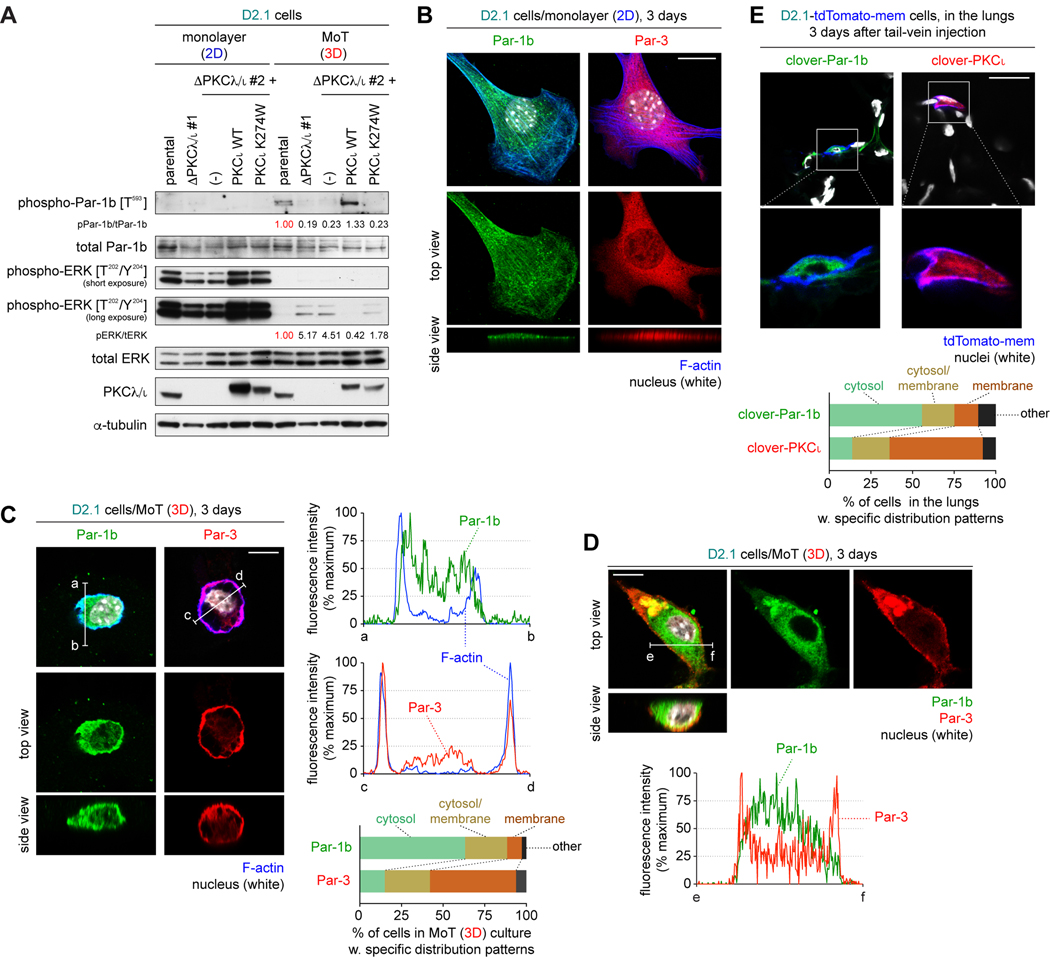
Systemic dissemination of tumor cells often begins long before the development of overt metastases, revealing the inefficient nature of the metastatic process. Thus, already at the time of initial clinical presentation, many patients with cancer harbor a myriad disseminated tumor cells (DTC) throughout the body, most of which are found as mitotically quiescent solitary cells. This indicates that the majority of DTCs fail, for still unknown reasons, to initiate rapid proliferation after entering foreign tissue, which likely contributes significantly to the inefficiency of metastasis formation. Here, we showed that extracellular matrix (ECM) components of the host parenchyma prevented proliferation of DTCs that had recently infiltrated foreign tissue by binding to syndecan receptors expressed on the surface of these cells. This led to the recruitment of the Par-3:Par-6:atypical PKC protein complex, a critical regulator of cell polarity, to the plasma membrane and release of Par-1 kinase into the cytosol. Cytosolic Par-1 bound, phosphorylated, and inactivated KSR scaffolding proteins ultimately inhibited Ras/ERK signaling and, in turn, cell proliferation. Inhibition of the syndecan-mediated signaling restored the proliferation of otherwise dormant DTCs, enabling these cells to efficiently colonize foreign tissues. Intriguingly, naturally aggressive cancer cells overcame the antiproliferative effect of syndecan-mediated signaling either by shutting down this signaling pathway or by activating a proproliferative signaling pathway that works independent of syndecan-mediated signaling. Collectively, these observations indicate that the proliferative arrest of DTCs is attributable, in part, to the syndecan-mediated ligation of ECM proteins. SIGNIFICANCE: This study identifies a novel signaling pathway that regulates the proliferative dormancy of individual disseminated tumor cells.Graphical Abstract: http://cancerres.aacrjournals.org/content/canres/79/23/5944/F1.large.jpg.
©2019 American Association for Cancer Research.
Conflict of interest statement
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Figures
Similar articles
-
Visualizing the Cell-Matrix Interactions and Cytoskeleton of Disseminated Tumor Cells.Methods Mol Biol. 2024;2811:207-220. doi: 10.1007/978-1-0716-3882-8_16. Methods Mol Biol. 2024. PMID: 39037661
-
Syndecans in heart fibrosis.Cell Tissue Res. 2016 Sep;365(3):539-52. doi: 10.1007/s00441-016-2454-2. Epub 2016 Jul 14. Cell Tissue Res. 2016. PMID: 27411689 Review.
-
A Feed-Forward Mechanosignaling Loop Confers Resistance to Therapies Targeting the MAPK Pathway in BRAF-Mutant Melanoma.Cancer Res. 2020 May 15;80(10):1927-1941. doi: 10.1158/0008-5472.CAN-19-2914. Epub 2020 Mar 16. Cancer Res. 2020. PMID: 32179513
-
Emerging Role of Syndecans in Extracellular Matrix Remodeling in Cancer.J Histochem Cytochem. 2020 Dec;68(12):863-870. doi: 10.1369/0022155420930112. Epub 2020 Jul 6. J Histochem Cytochem. 2020. PMID: 32623937 Free PMC article. Review.
-
RASSF1A Deficiency Enhances RAS-Driven Lung Tumorigenesis.Cancer Res. 2018 May 15;78(10):2614-2623. doi: 10.1158/0008-5472.CAN-17-2466. Epub 2018 May 7. Cancer Res. 2018. PMID: 29735543 Free PMC article.
Cited by
-
Extracellular matrix remodeling in tumor progression and immune escape: from mechanisms to treatments.Mol Cancer. 2023 Mar 11;22(1):48. doi: 10.1186/s12943-023-01744-8. Mol Cancer. 2023. PMID: 36906534 Free PMC article. Review.
-
Rivaroxaban as a Protector of Oxidative Stress-Induced Vascular Endothelial Glycocalyx Damage via the IQGAP1/PAR1-2/PI3K/Akt Pathway.J Vasc Res. 2025;62(1):22-36. doi: 10.1159/000542419. Epub 2024 Nov 2. J Vasc Res. 2025. PMID: 39496251 Free PMC article.
-
Visualizing the Cell-Matrix Interactions and Cytoskeleton of Disseminated Tumor Cells.Methods Mol Biol. 2024;2811:207-220. doi: 10.1007/978-1-0716-3882-8_16. Methods Mol Biol. 2024. PMID: 39037661
-
Syndecan-1 (CD138), Carcinomas and EMT.Int J Mol Sci. 2021 Apr 19;22(8):4227. doi: 10.3390/ijms22084227. Int J Mol Sci. 2021. PMID: 33921767 Free PMC article. Review.
-
Sustained Oncogenic Signaling in the Cytostatic State Enables Targeting of Nonproliferating Persistent Cancer Cells.Cancer Res. 2022 Sep 2;82(17):3045-3057. doi: 10.1158/0008-5472.CAN-21-2908. Cancer Res. 2022. PMID: 35792658 Free PMC article.
References
-
- Naumov GN, MacDonald IC, Weinmeister PM, Kerkvliet N, Nadkarni KV, Wilson SM, et al. Persistence of solitary mammary carcinoma cells in a secondary site: a possible contributor to dormancy. Cancer Res 2002;62(7):2162–8. - PubMed
-
- Pantel K, Alix-Panabieres C, Riethdorf S. Cancer micrometastases. Nat Rev Clin Oncol 2009;6(6):339–51. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
