

Host-transposon interactions: conflict, cooperation, and cooption
- PMID: 31481535
- PMCID: PMC6719617
- DOI: 10.1101/gad.327312.119
Host-transposon interactions: conflict, cooperation, and cooption
Abstract
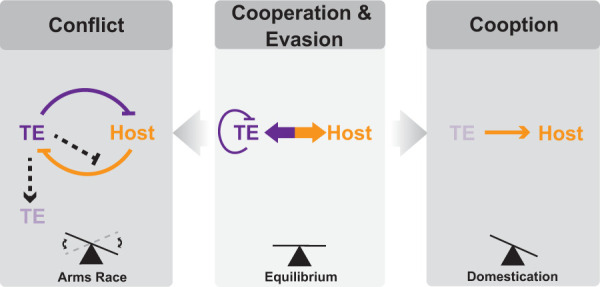
Transposable elements (TEs) are mobile DNA sequences that colonize genomes and threaten genome integrity. As a result, several mechanisms appear to have emerged during eukaryotic evolution to suppress TE activity. However, TEs are ubiquitous and account for a prominent fraction of most eukaryotic genomes. We argue that the evolutionary success of TEs cannot be explained solely by evasion from host control mechanisms. Rather, some TEs have evolved commensal and even mutualistic strategies that mitigate the cost of their propagation. These coevolutionary processes promote the emergence of complex cellular activities, which in turn pave the way for cooption of TE sequences for organismal function.
Keywords: KRAB zinc finger; fetrotransposons; gene regulation; genomics; piRNA; transposable elements.
© 2019 Cosby et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
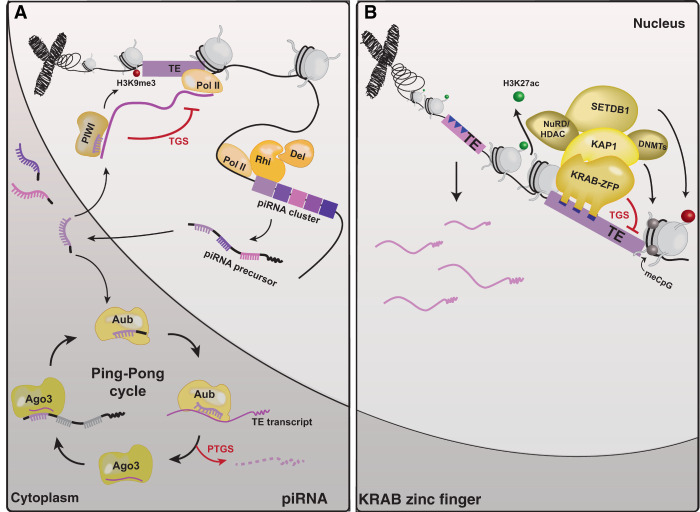
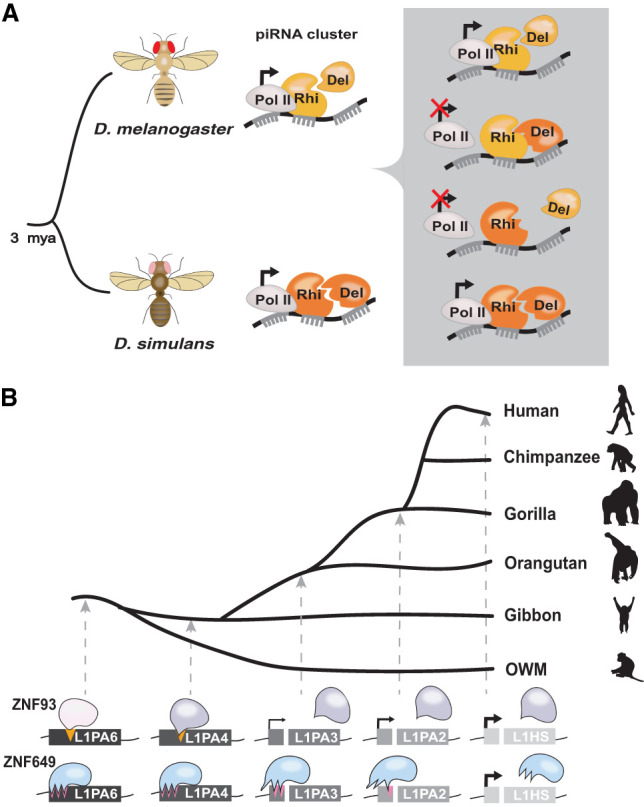
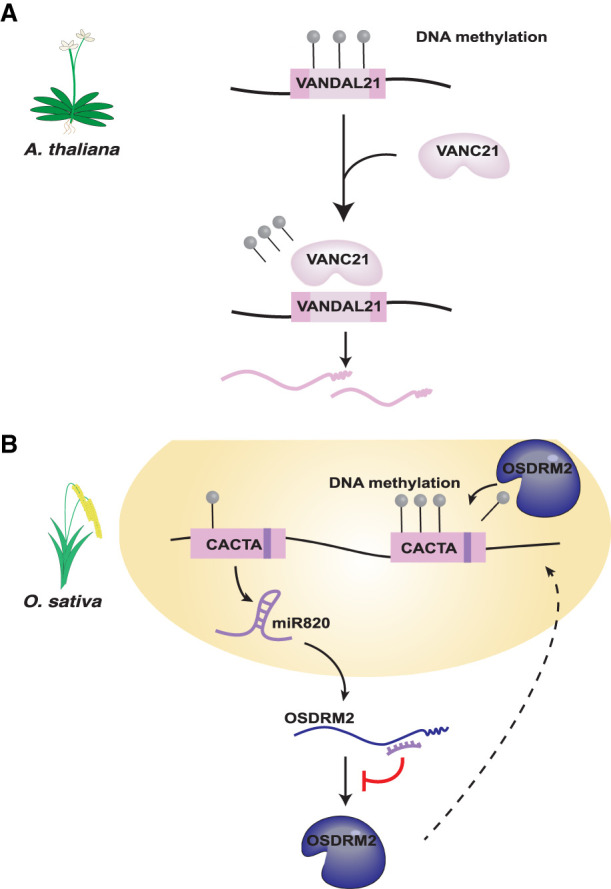
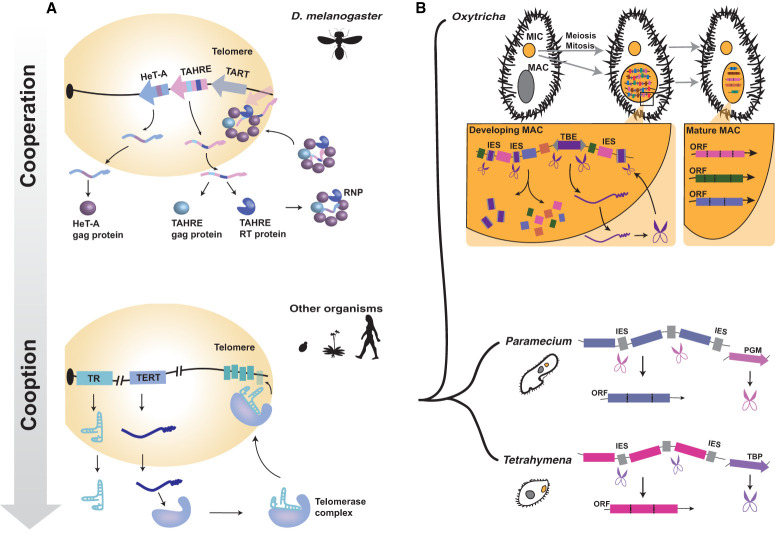
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials