

Microtubules gate tau condensation to spatially regulate microtubule functions
- PMID: 31481790
- PMCID: PMC6748660
- DOI: 10.1038/s41556-019-0375-5
Microtubules gate tau condensation to spatially regulate microtubule functions
Abstract
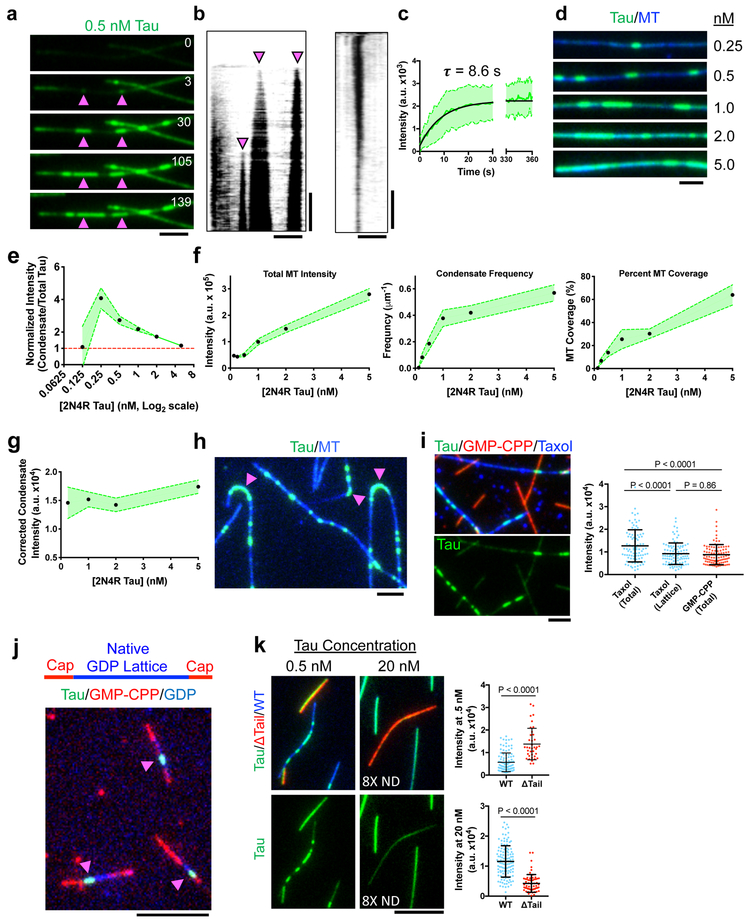
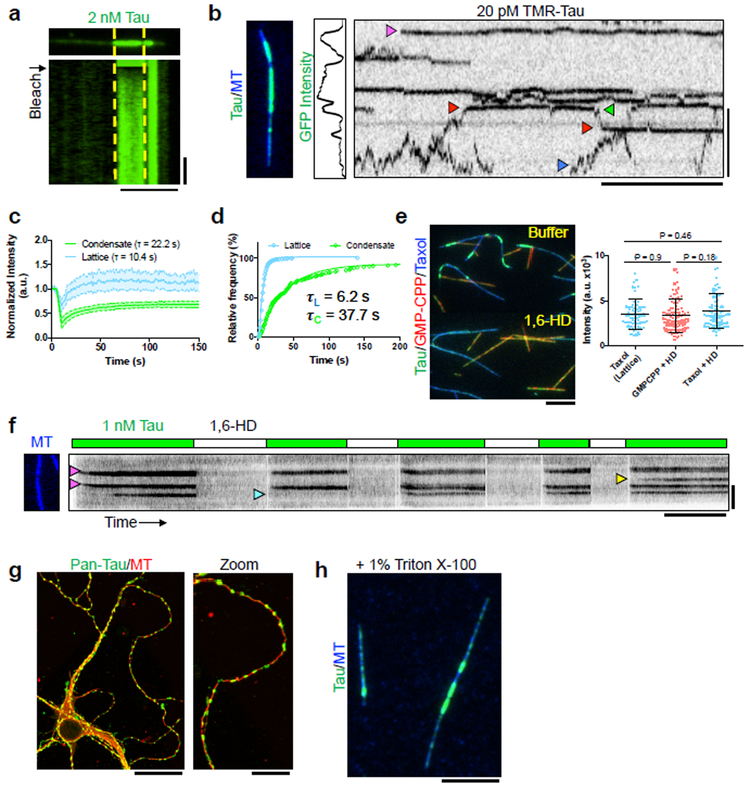
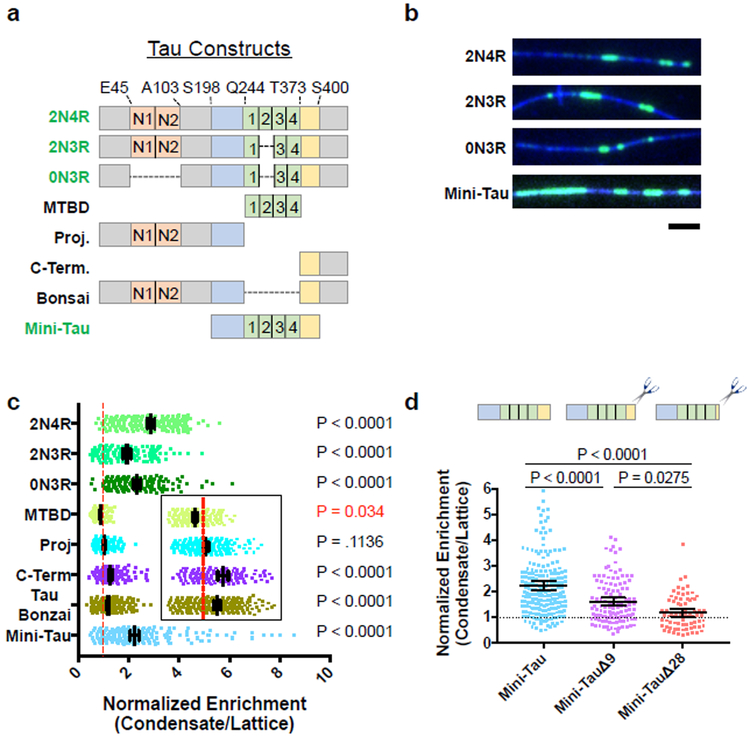
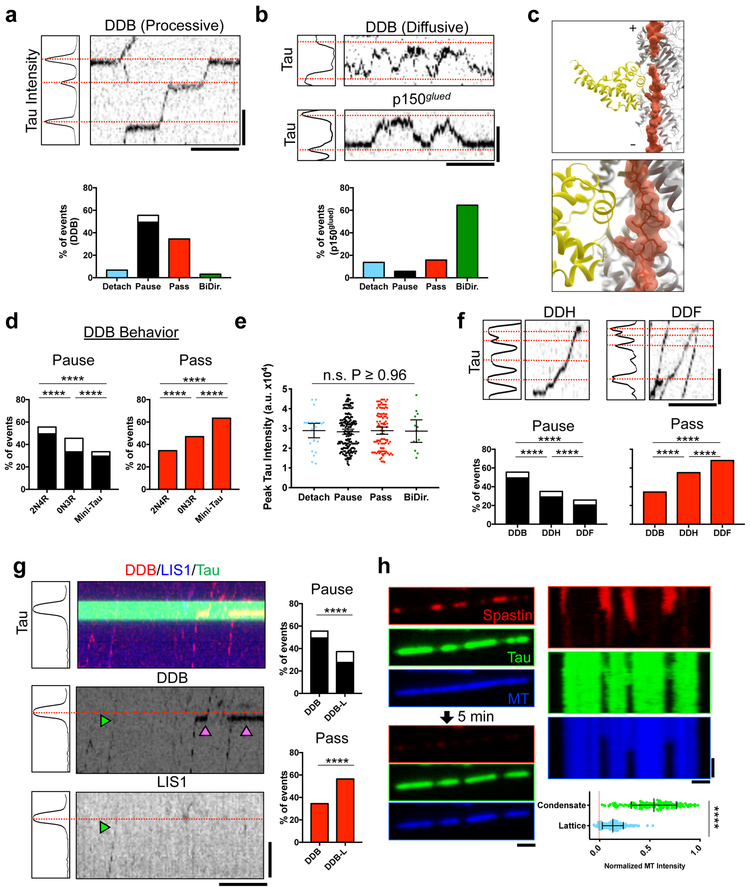
Tau is an abundant microtubule-associated protein in neurons. Tau aggregation into insoluble fibrils is a hallmark of Alzheimer's disease and other types of dementia1, yet the physiological state of tau molecules within cells remains unclear. Using single-molecule imaging, we directly observe that the microtubule lattice regulates reversible tau self-association, leading to localized, dynamic condensation of tau molecules on the microtubule surface. Tau condensates form selectively permissible barriers, spatially regulating the activity of microtubule-severing enzymes and the movement of molecular motors through their boundaries. We propose that reversible self-association of tau molecules, gated by the microtubule lattice, is an important mechanism of the biological functions of tau, and that oligomerization of tau is a common property shared between the physiological and disease-associated forms of the molecule.
Conflict of interest statement
Declaration of Interests
Authors declare no financial and non-financial competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
