

Spontaneous Isomerization of Long-Lived Proteins Provides a Molecular Mechanism for the Lysosomal Failure Observed in Alzheimer's Disease
- PMID: 31482121
- PMCID: PMC6716341
- DOI: 10.1021/acscentsci.9b00369
Spontaneous Isomerization of Long-Lived Proteins Provides a Molecular Mechanism for the Lysosomal Failure Observed in Alzheimer's Disease
Abstract
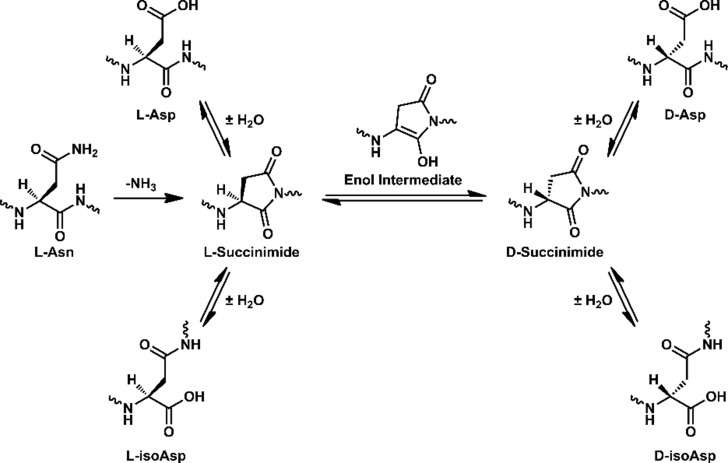
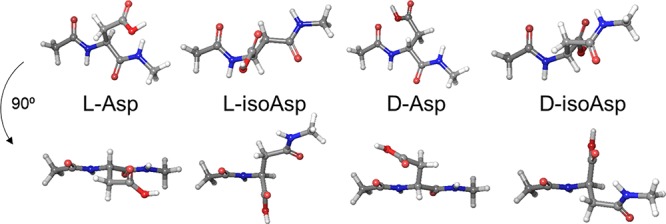
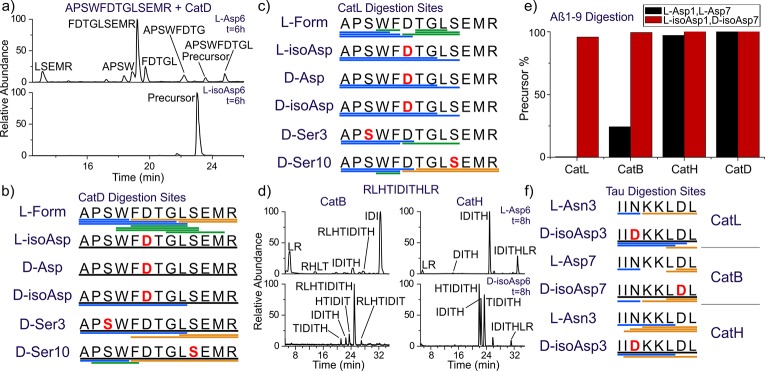
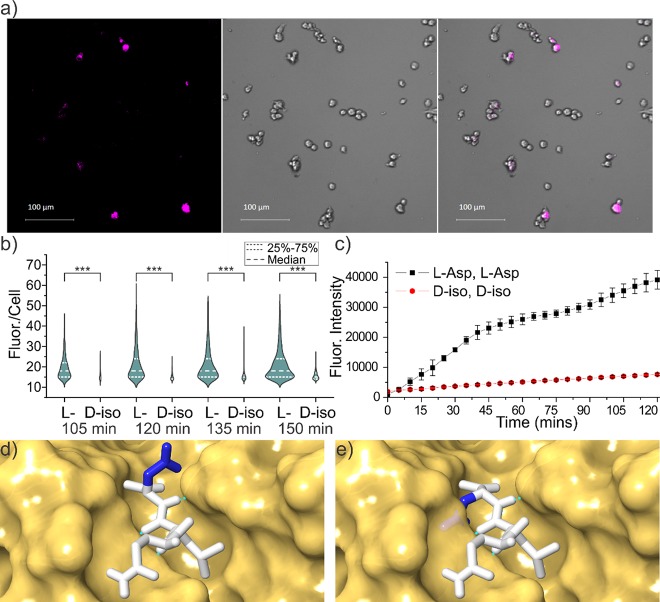
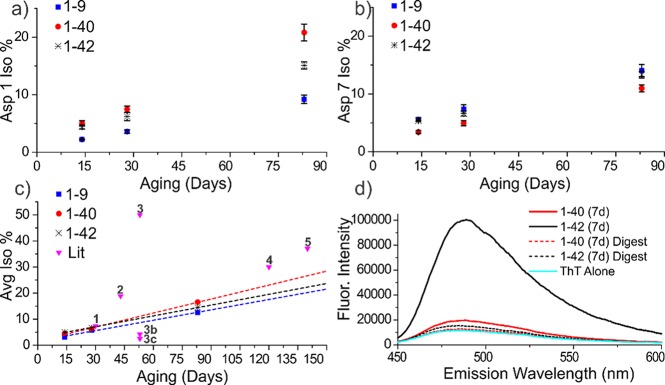
Proteinaceous aggregation is a well-known observable in Alzheimer's disease (AD), but failure and storage of lysosomal bodies within neurons is equally ubiquitous and actually precedes bulk accumulation of extracellular amyloid plaque. In fact, AD shares many similarities with certain lysosomal storage disorders though establishing a biochemical connection has proven difficult. Herein, we demonstrate that isomerization and epimerization, which are spontaneous chemical modifications that occur in long-lived proteins, prevent digestion by the proteases in the lysosome (namely, the cathepsins). For example, isomerization of aspartic acid into l-isoAsp prevents digestion of the N-terminal portion of Aβ by cathepsin L, one of the most aggressive lysosomal proteases. Similar results were obtained after examination of various target peptides with a full series of cathepsins, including endo-, amino-, and carboxy-peptidases. In all cases peptide fragments too long for transporter recognition or release from the lysosome persisted after treatment, providing a mechanism for eventual lysosomal storage and bridging the gap between AD and lysosomal storage disorders. Additional experiments with microglial cells confirmed that isomerization disrupts proteolysis in active lysosomes. These results are easily rationalized in terms of protease active sites, which are engineered to precisely orient the peptide backbone and cannot accommodate the backbone shift caused by isoaspartic acid or side chain dislocation resulting from epimerization. Although Aβ is known to be isomerized and epimerized in plaques present in AD brains, we further establish that the rates of modification for aspartic acid in positions 1 and 7 are fast and could accrue prior to plaque formation. Spontaneous chemistry can therefore provide modified substrates capable of inducing gradual lysosomal failure, which may play an important role in the cascade of events leading to the disrupted proteostasis, amyloid formation, and tauopathies associated with AD.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
