

Cold adaptation in the environmental bacterium Shewanella oneidensis is controlled by a J-domain co-chaperone protein network
- PMID: 31482142
- PMCID: PMC6715715
- DOI: 10.1038/s42003-019-0567-3
Cold adaptation in the environmental bacterium Shewanella oneidensis is controlled by a J-domain co-chaperone protein network
Abstract
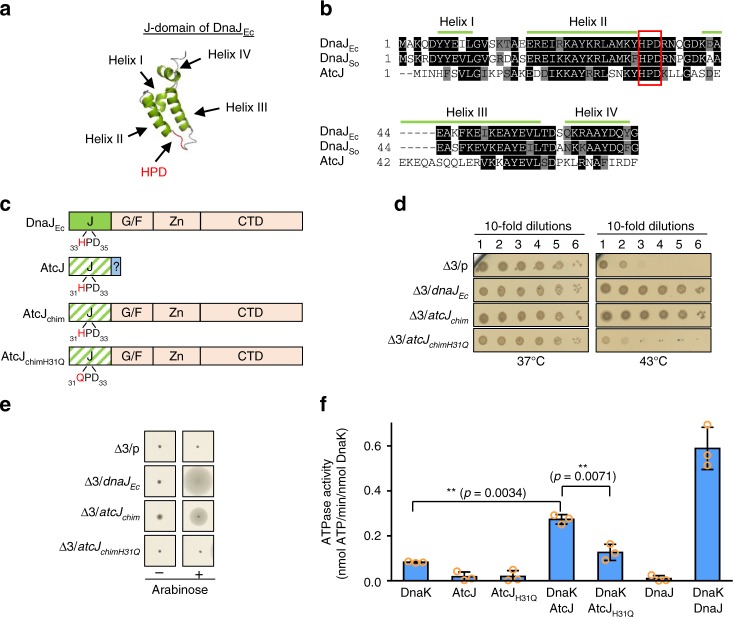
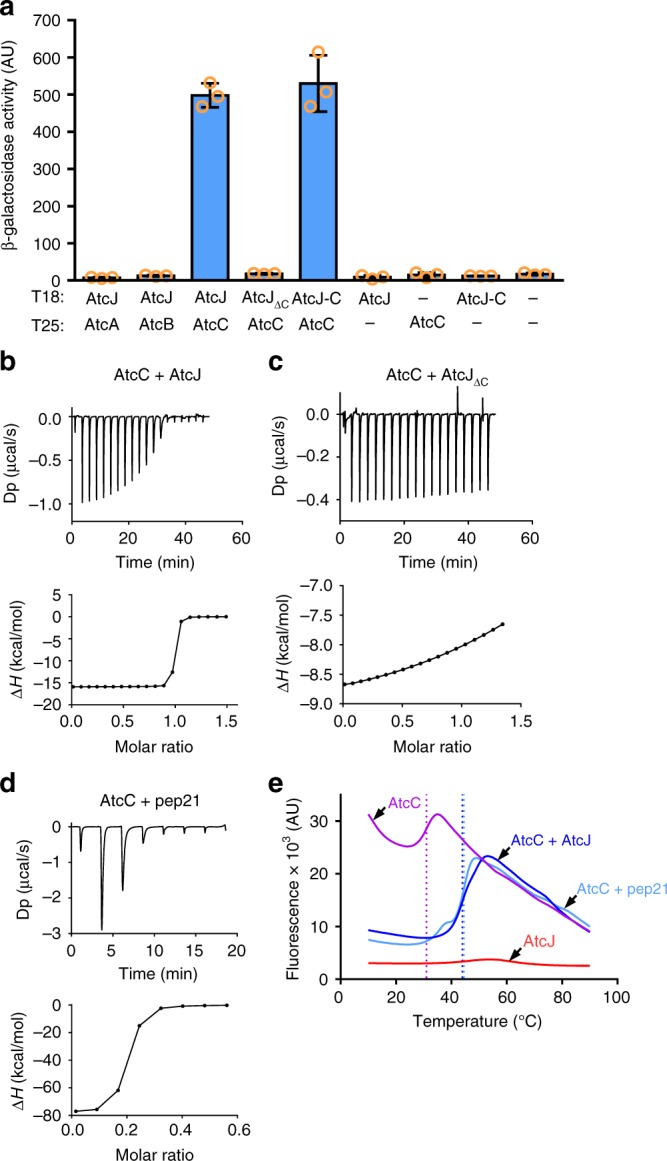
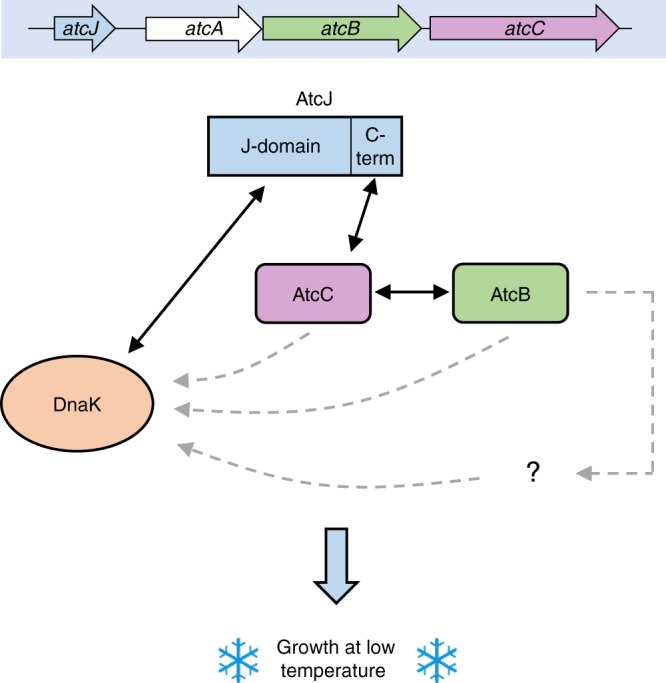
DnaK (Hsp70) is a major ATP-dependent chaperone that functions with two co-chaperones, a J-domain protein (JDP) and a nucleotide exchange factor to maintain proteostasis in most organisms. Here, we show that the environmental bacterium Shewanella oneidensis possesses a previously uncharacterized short JDP, AtcJ, dedicated to cold adaptation and composed of a functional J-domain and a C-terminal extension of 21 amino acids. We showed that atcJ is the first gene of an operon encoding also AtcA, AtcB and AtcC, three proteins of unknown functions. Interestingly, we found that the absence of AtcJ, AtcB or AtcC leads to a dramatically reduced growth at low temperature. In addition, we demonstrated that AtcJ interacts via its C-terminal extension with AtcC, and that AtcC binds to AtcB. Therefore, we identified a previously uncharacterized protein network that involves the DnaK system with a dedicated JDP to allow bacteria to survive to cold environment.
Keywords: Bacterial physiology; Chaperones.
Conflict of interest statement
Competing interestsThe authors declare no competing interests.
Figures



References
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
