

Strong preference for autaptic self-connectivity of neocortical PV interneurons facilitates their tuning to γ-oscillations
- PMID: 31483783
- PMCID: PMC6726197
- DOI: 10.1371/journal.pbio.3000419
Strong preference for autaptic self-connectivity of neocortical PV interneurons facilitates their tuning to γ-oscillations
Abstract
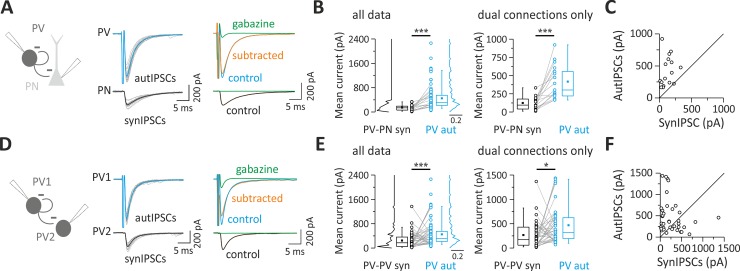
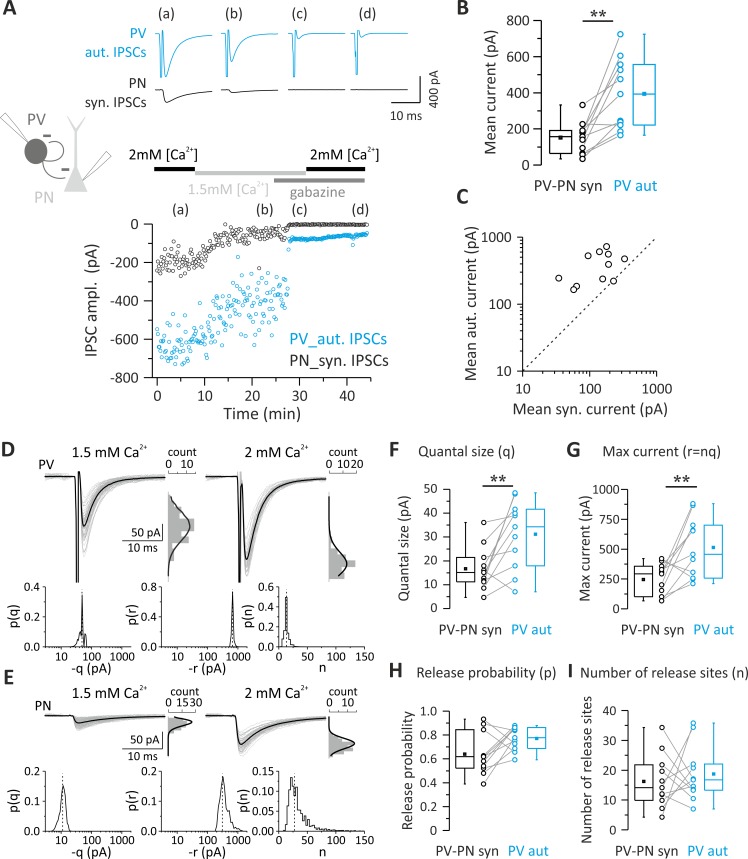
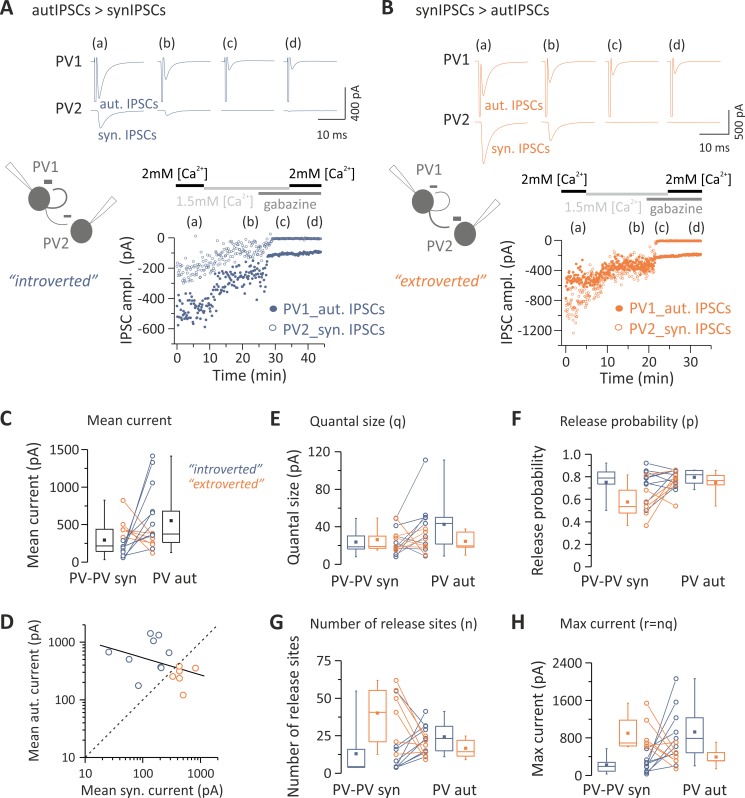
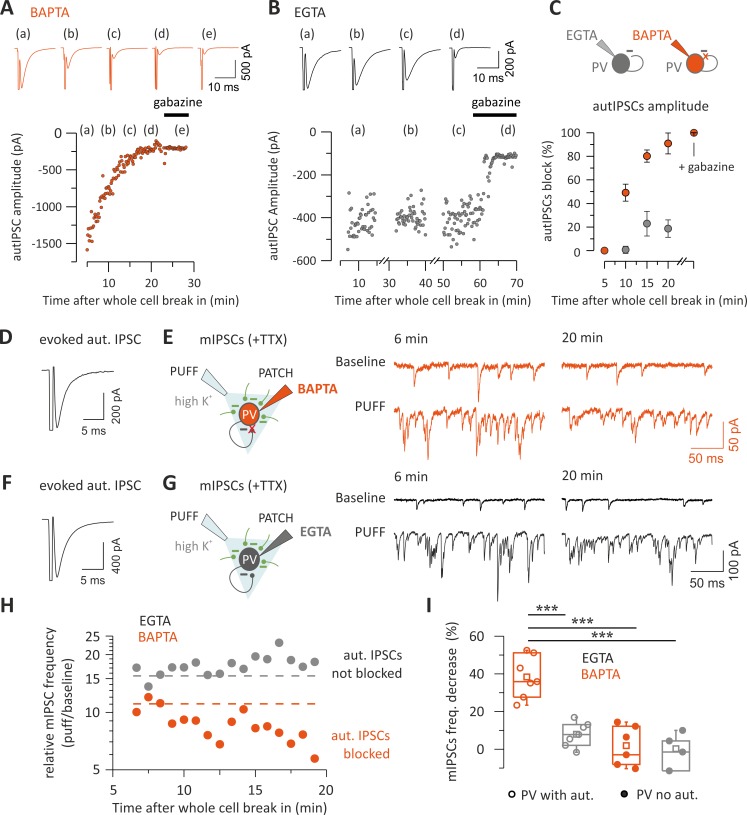
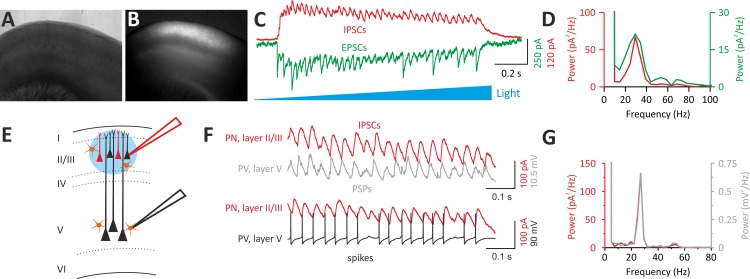
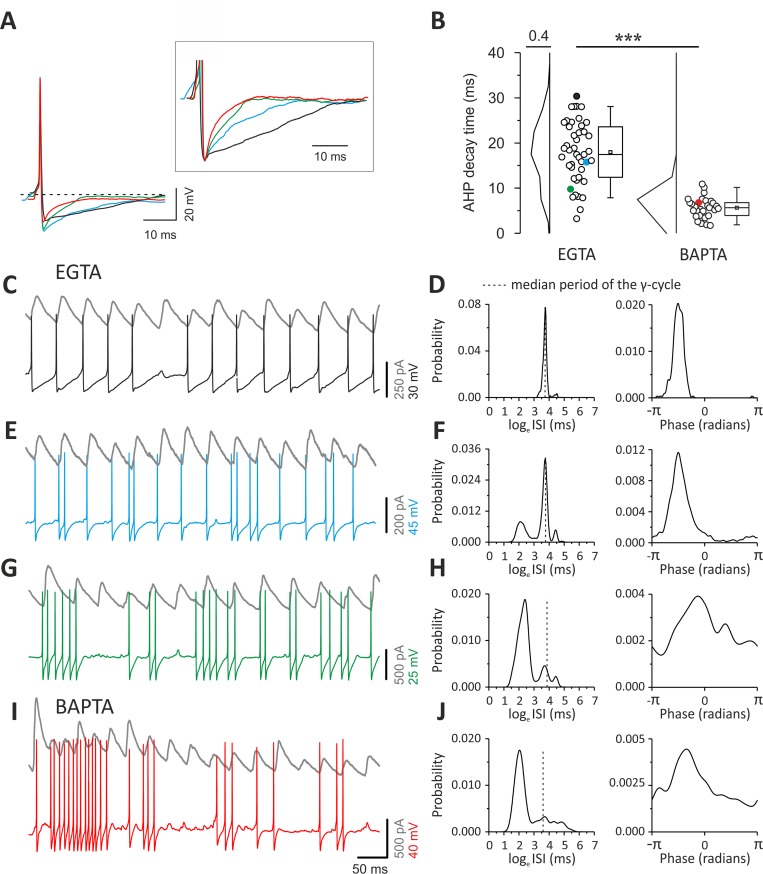
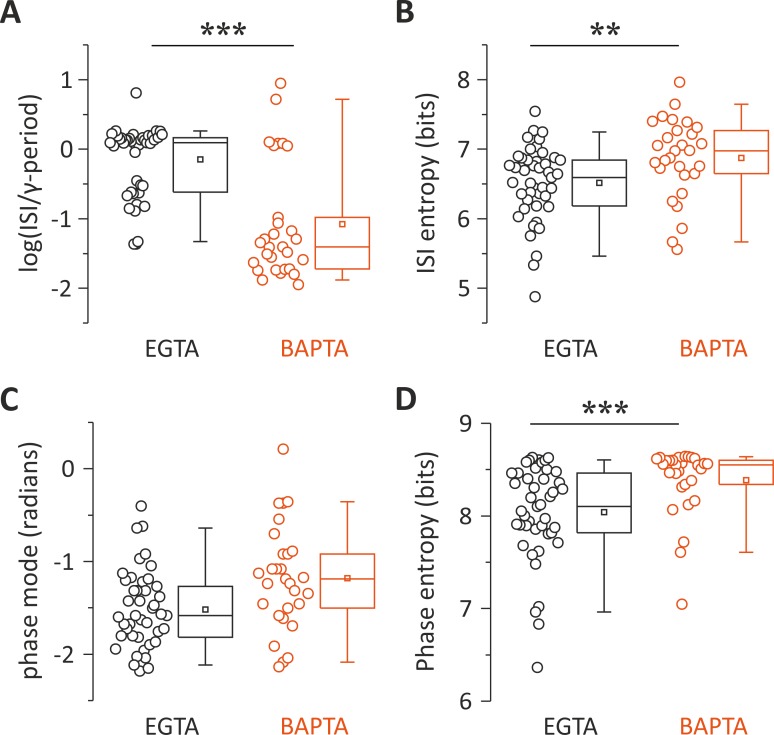
Parvalbumin (PV)-positive interneurons modulate cortical activity through highly specialized connectivity patterns onto excitatory pyramidal neurons (PNs) and other inhibitory cells. PV cells are autoconnected through powerful autapses, but the contribution of this form of fast disinhibition to cortical function is unknown. We found that autaptic transmission represents the most powerful inhibitory input of PV cells in neocortical layer V. Autaptic strength was greater than synaptic strength onto PNs as a result of a larger quantal size, whereas autaptic and heterosynaptic PV-PV synapses differed in the number of release sites. Overall, single-axon autaptic transmission contributed to approximately 40% of the global inhibition (mostly perisomatic) that PV interneurons received. The strength of autaptic transmission modulated the coupling of PV-cell firing with optogenetically induced γ-oscillations, preventing high-frequency bursts of spikes. Autaptic self-inhibition represents an exceptionally large and fast disinhibitory mechanism, favoring synchronization of PV-cell firing during cognitive-relevant cortical network activity.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
