

HilD induces expression of a novel Salmonella Typhimurium invasion factor, YobH, through a regulatory cascade involving SprB
- PMID: 31484980
- PMCID: PMC6726612
- DOI: 10.1038/s41598-019-49192-z
HilD induces expression of a novel Salmonella Typhimurium invasion factor, YobH, through a regulatory cascade involving SprB
Abstract
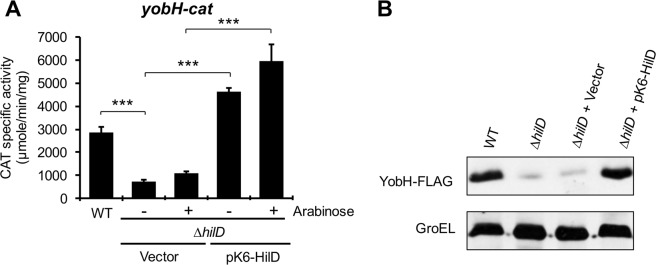
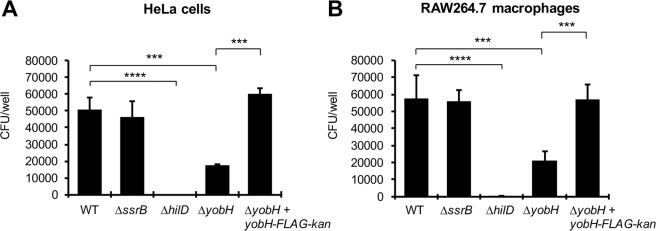
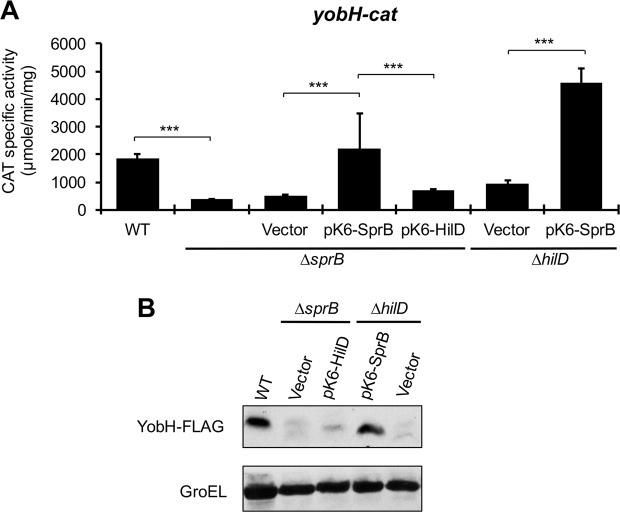
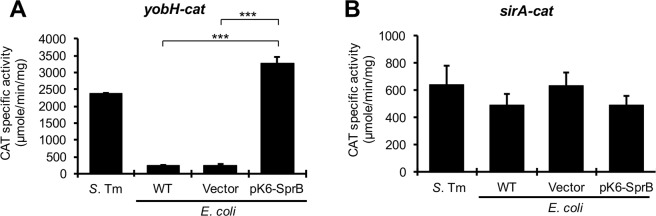
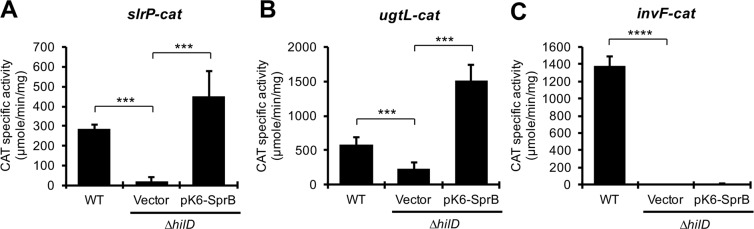
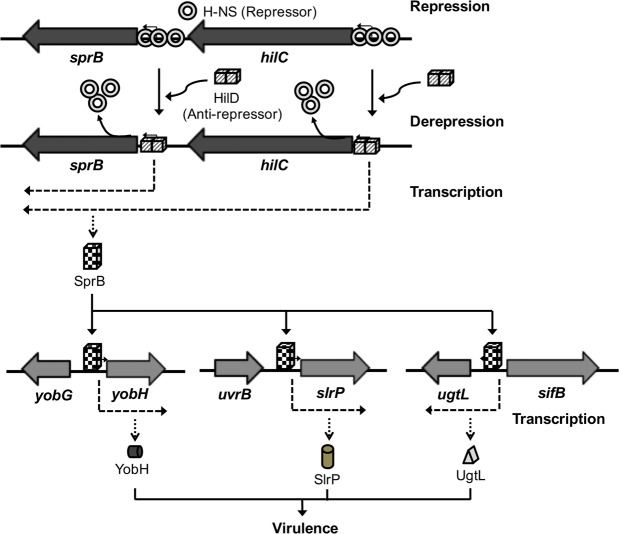
HilD is an AraC-like transcriptional regulator encoded in the Salmonella pathogenicity island 1 (SPI-1), which actives transcription of many genes within and outside SPI-1 that are mainly required for invasion of Salmonella into host cells. HilD controls expression of target genes directly or by acting through distinct regulators; three different regulatory cascades headed by HilD have been described to date. Here, by analyzing the effect of HilD on the yobH gene in Salmonella enterica serovar Typhimurium (S. Typhimurium), we further define an additional regulatory cascade mediated by HilD, which was revealed by previous genome-wide analyses. In this regulatory cascade, HilD acts through SprB, a LuxR-like regulator encoded in SPI-1, to induce expression of virulence genes. Our data show that HilD induces expression of sprB by directly counteracting H-NS-mediated repression on the promoter region upstream of this gene. Then, SprB directly activates expression of several genes including yobH, slrP and ugtL. Interestingly, we found that YobH, a protein of only 79 amino acids, is required for invasion of S. Typhimurium into HeLa cells and mouse macrophages. Thus, our results reveal a novel S. Typhimurium invasion factor and provide more evidence supporting the HilD-SprB regulatory cascade.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Eng S-K, et al. Salmonella: A review on pathogenesis, epidemiology and antibiotic resistance. Front. Life Sci. 2015;8:284–293. doi: 10.1080/21553769.2015.1051243. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
