

Maternal vitamin C regulates reprogramming of DNA methylation and germline development
- PMID: 31485074
- PMCID: PMC8423347
- DOI: 10.1038/s41586-019-1536-1
Maternal vitamin C regulates reprogramming of DNA methylation and germline development
Erratum in
-
Author Correction: Maternal vitamin C regulates reprogramming of DNA methylation and germline development.Nature. 2019 Dec;576(7785):E1. doi: 10.1038/s41586-019-1699-9. Nature. 2019. PMID: 31719689
Abstract
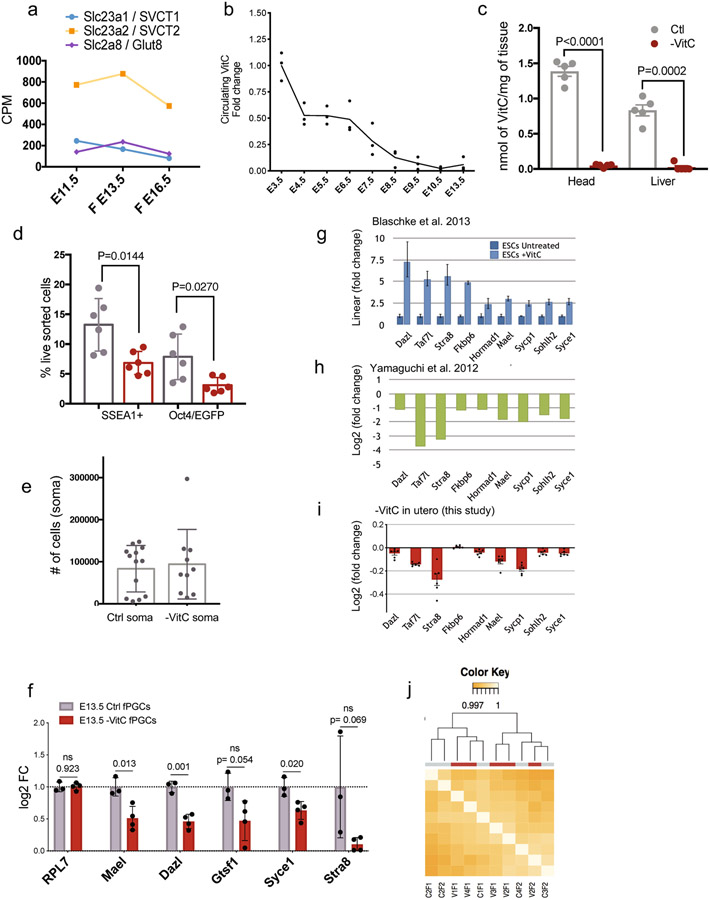
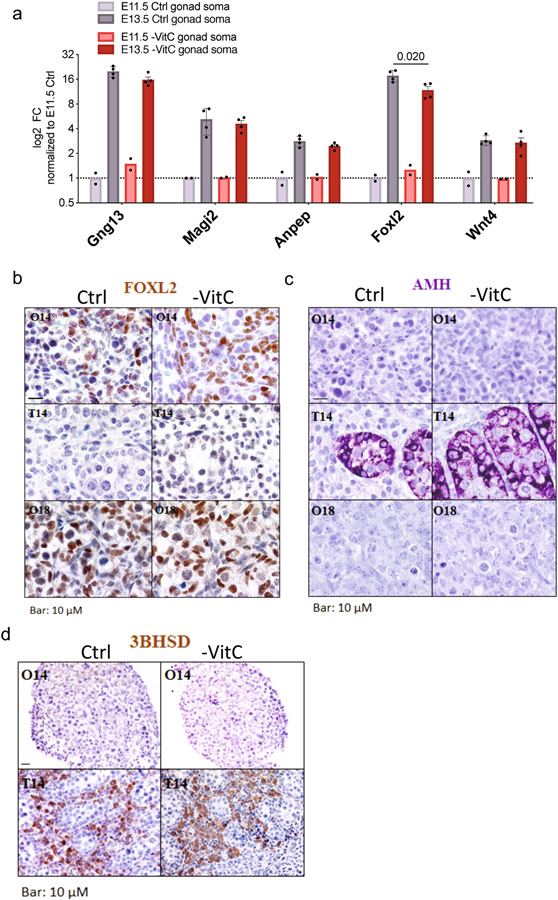
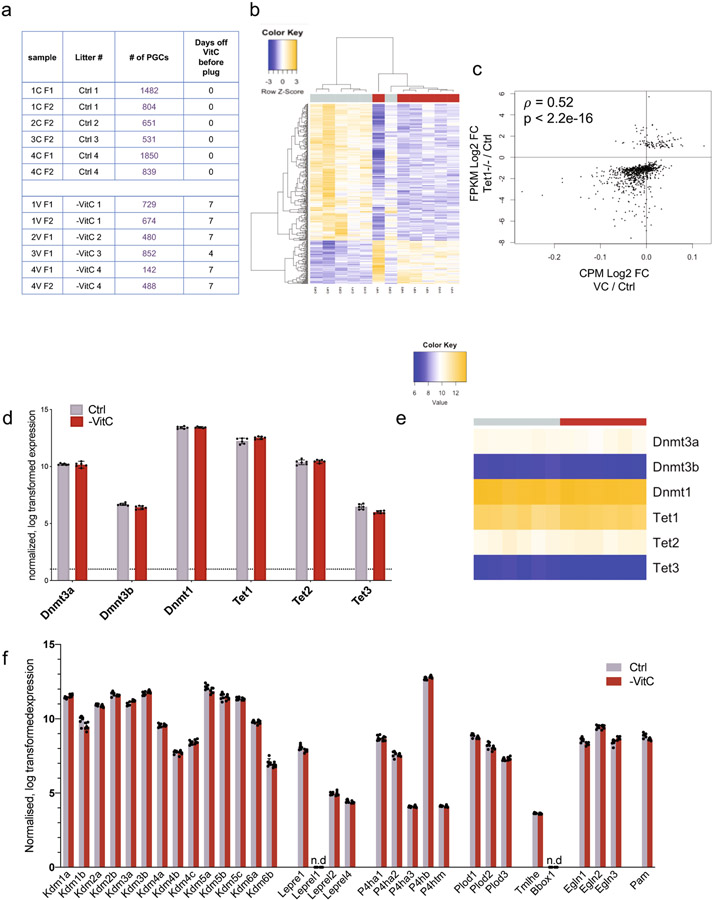
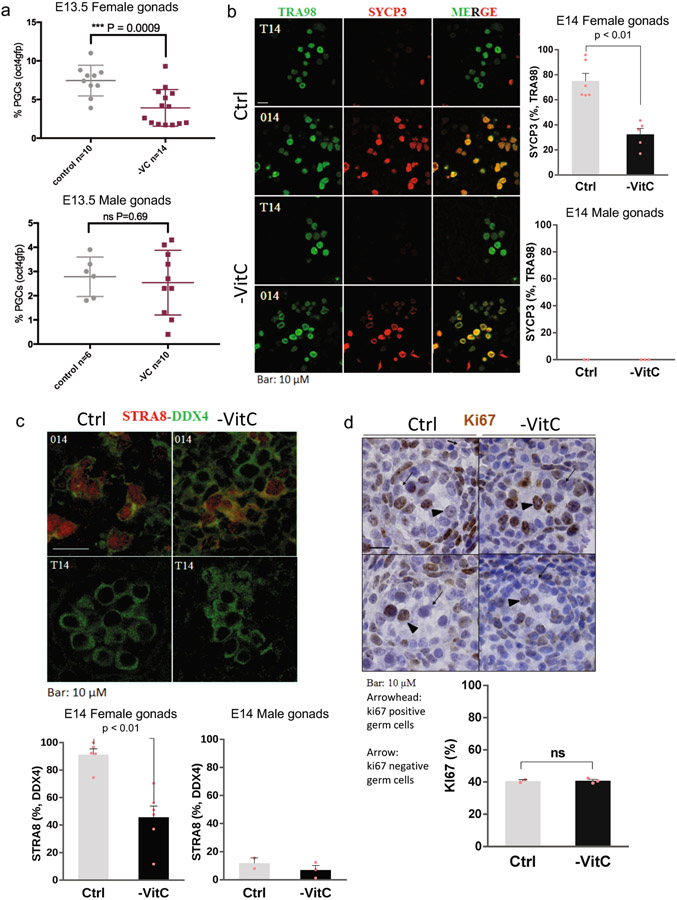
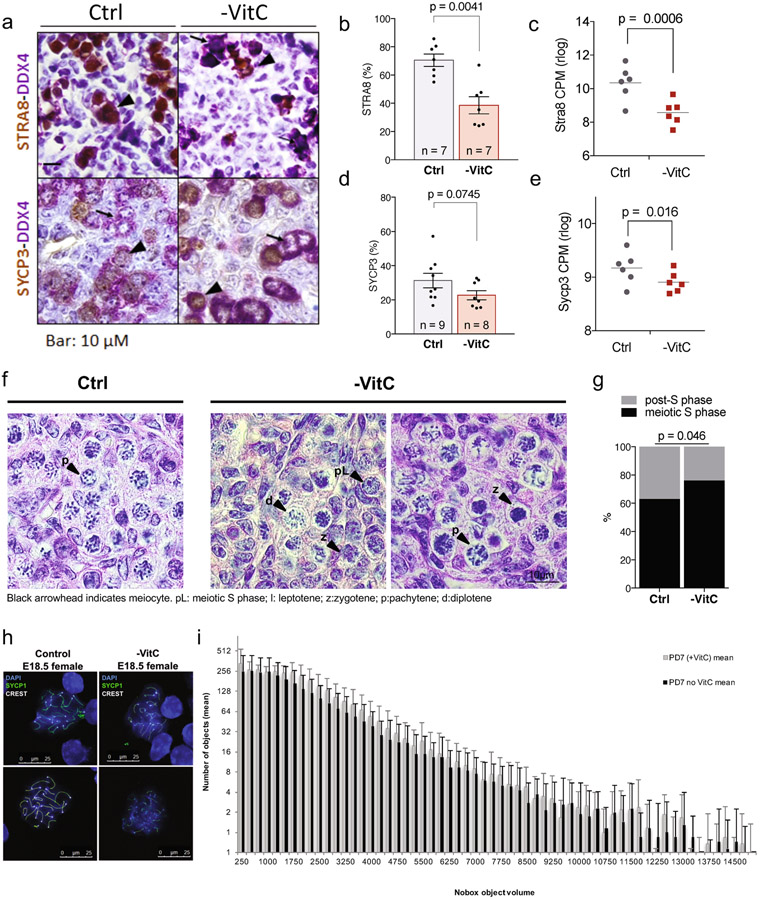
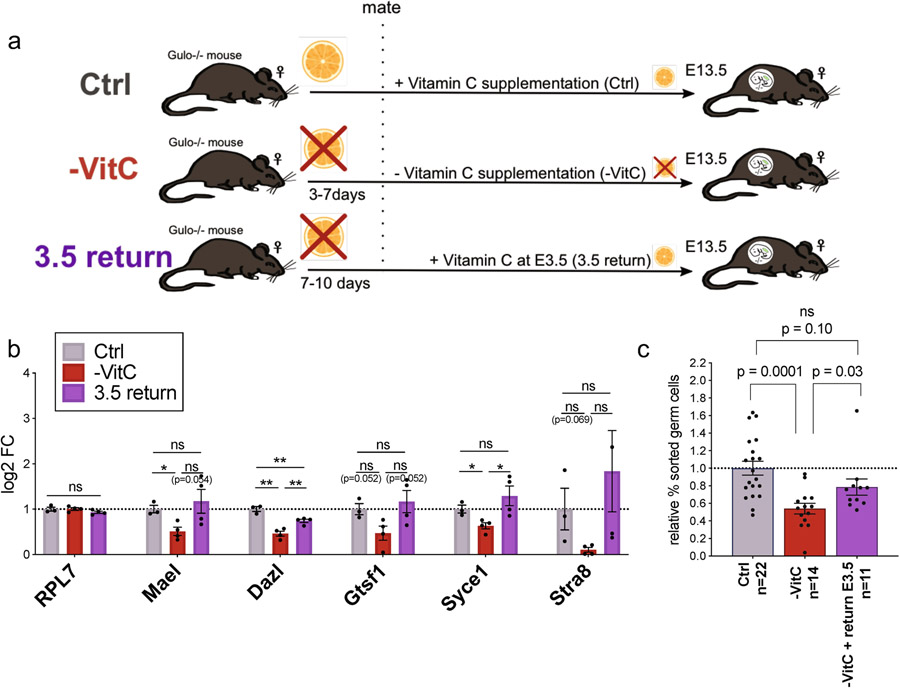
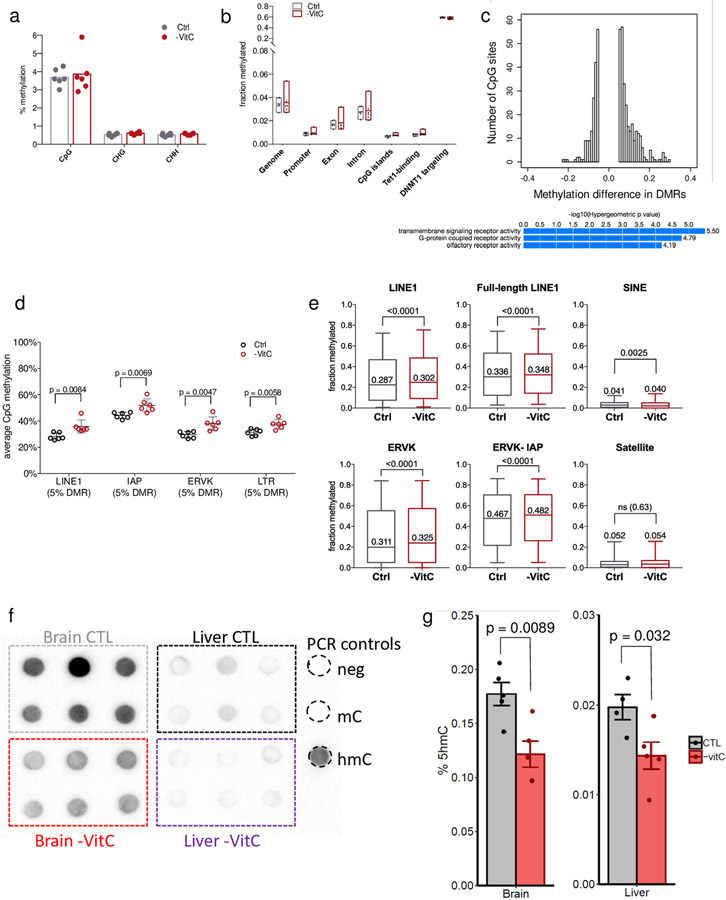
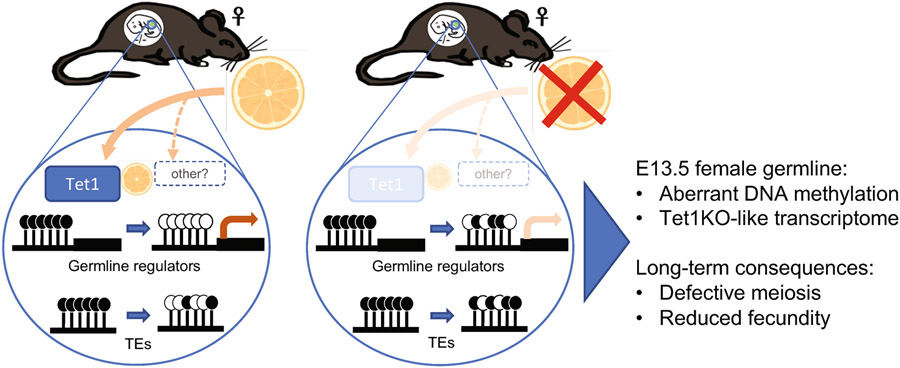
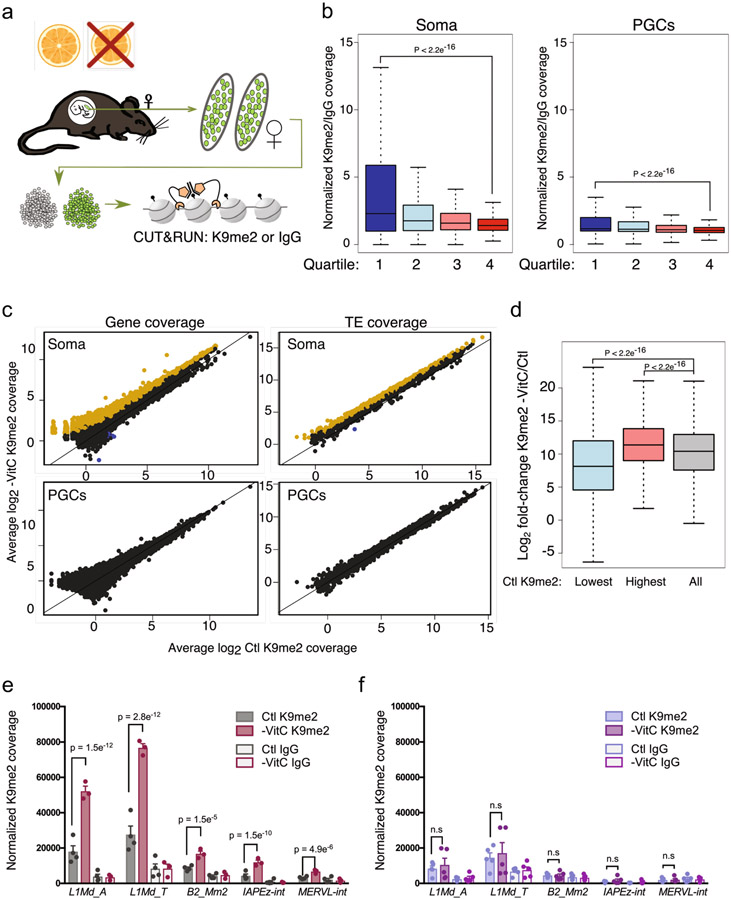
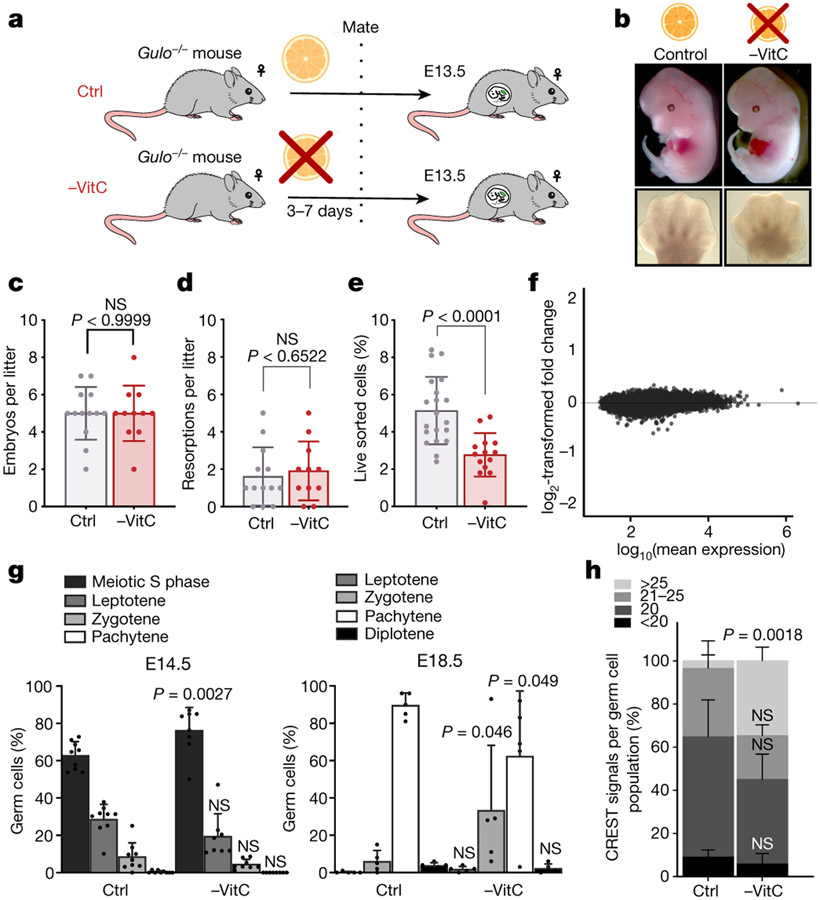
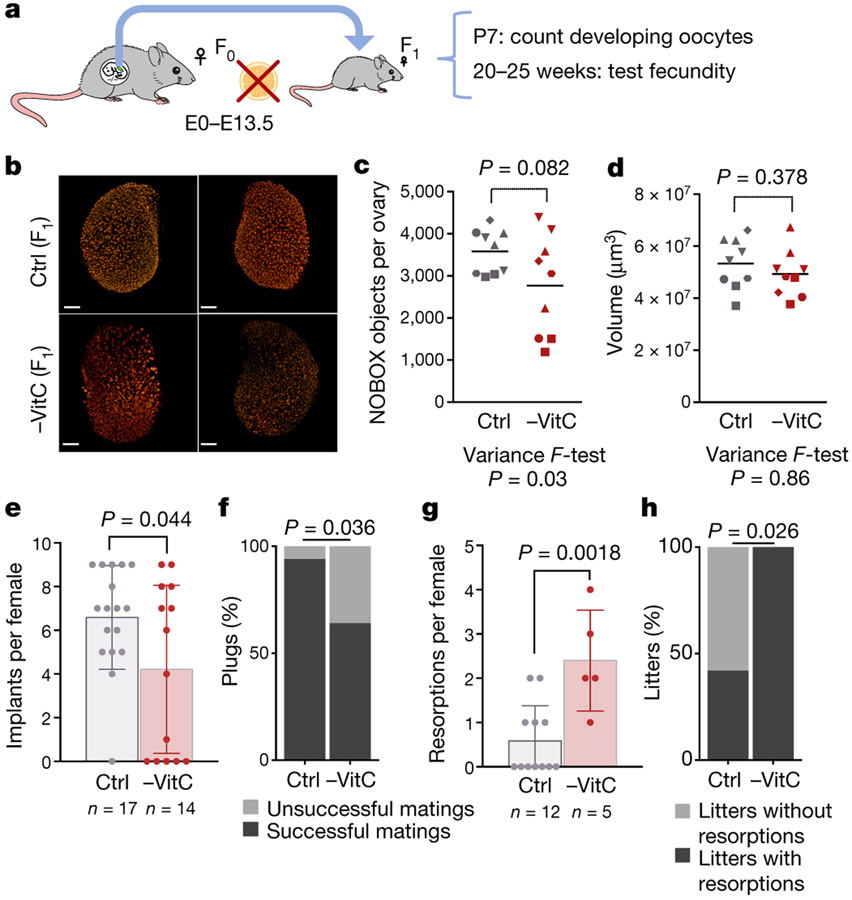
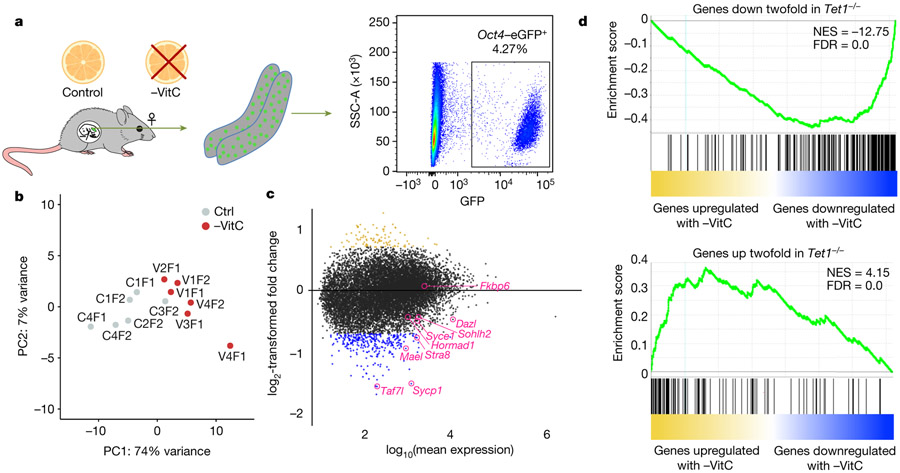
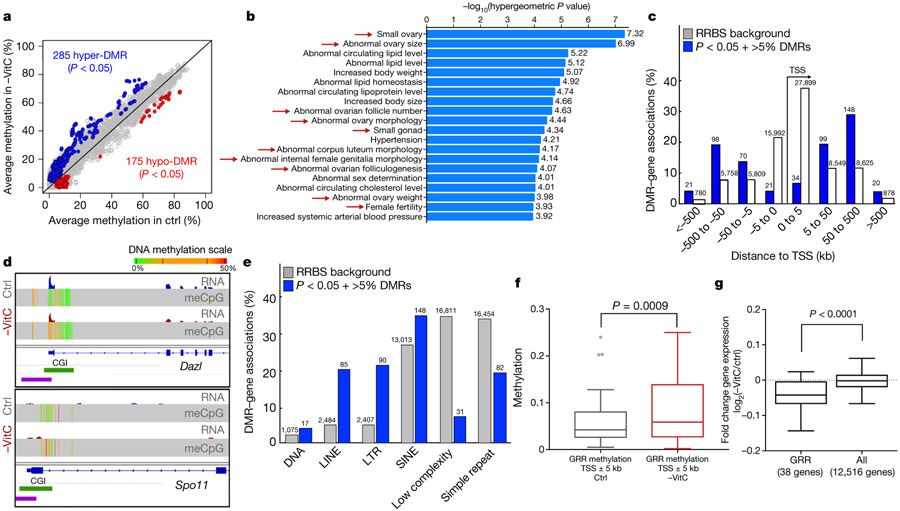
Development is often assumed to be hardwired in the genome, but several lines of evidence indicate that it is susceptible to environmental modulation with potential long-term consequences, including in mammals1,2. The embryonic germline is of particular interest because of the potential for intergenerational epigenetic effects. The mammalian germline undergoes extensive DNA demethylation3-7 that occurs in large part by passive dilution of methylation over successive cell divisions, accompanied by active DNA demethylation by TET enzymes3,8-10. TET activity has been shown to be modulated by nutrients and metabolites, such as vitamin C11-15. Here we show that maternal vitamin C is required for proper DNA demethylation and the development of female fetal germ cells in a mouse model. Maternal vitamin C deficiency does not affect overall embryonic development but leads to reduced numbers of germ cells, delayed meiosis and reduced fecundity in adult offspring. The transcriptome of germ cells from vitamin-C-deficient embryos is remarkably similar to that of embryos carrying a null mutation in Tet1. Vitamin C deficiency leads to an aberrant DNA methylation profile that includes incomplete demethylation of key regulators of meiosis and transposable elements. These findings reveal that deficiency in vitamin C during gestation partially recapitulates loss of TET1, and provide a potential intergenerational mechanism for adjusting fecundity to environmental conditions.
Figures
References
-
- Barker DJP The developmental origins of adult disease. J Am Coll Nutr 23, 588S–595S (2004). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
