

Structures of influenza A virus RNA polymerase offer insight into viral genome replication
- PMID: 31485076
- PMCID: PMC6795553
- DOI: 10.1038/s41586-019-1530-7
Structures of influenza A virus RNA polymerase offer insight into viral genome replication
Abstract
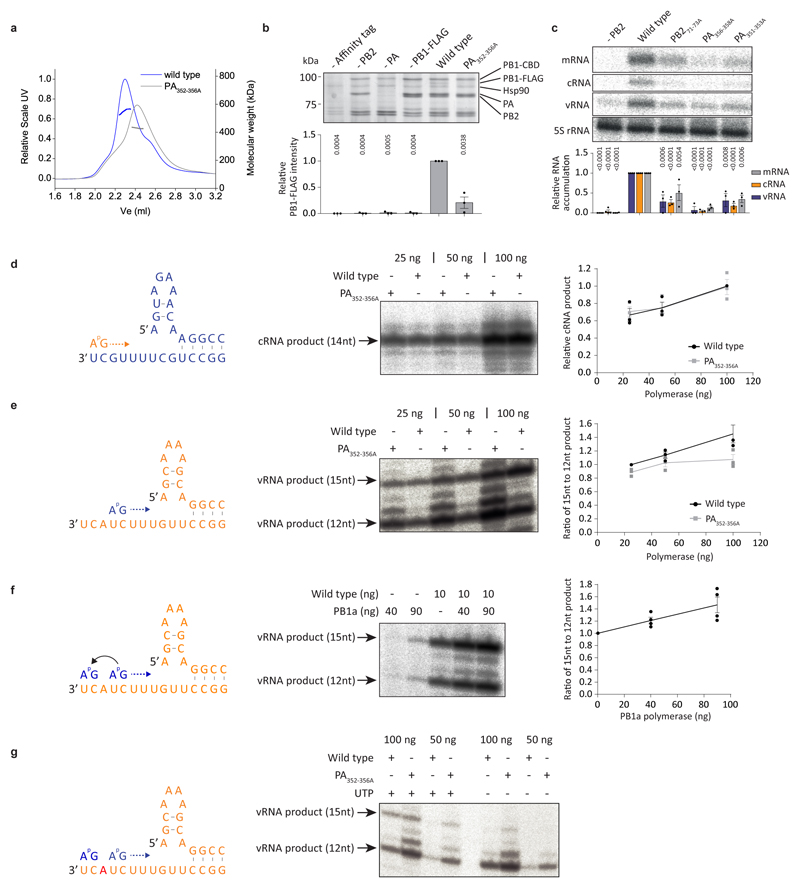
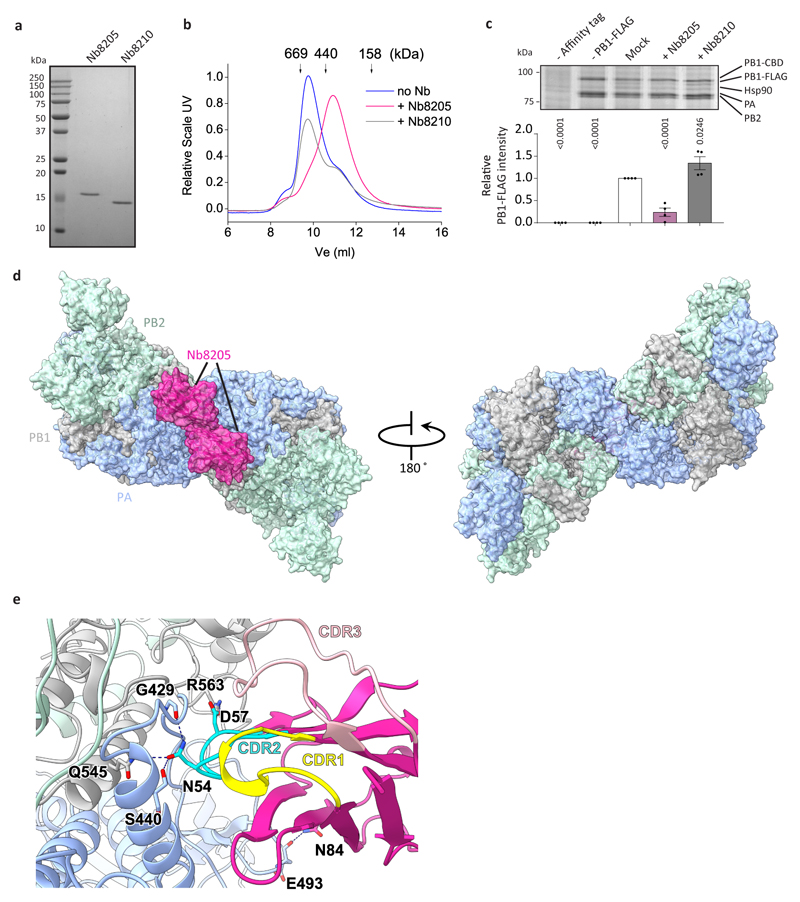
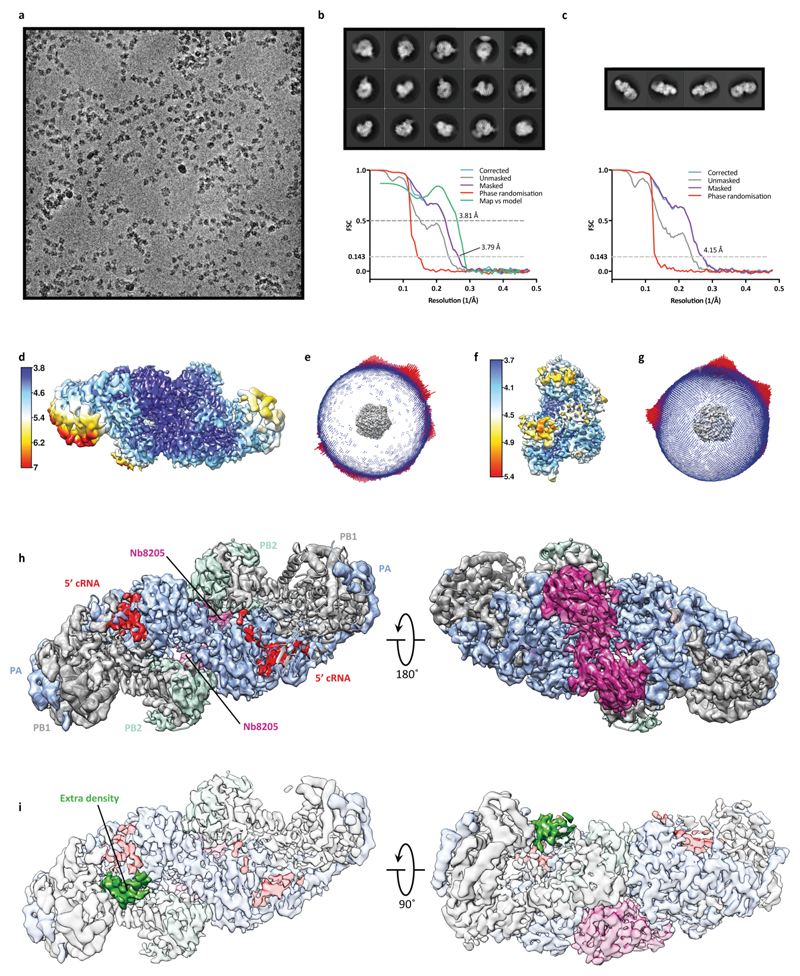
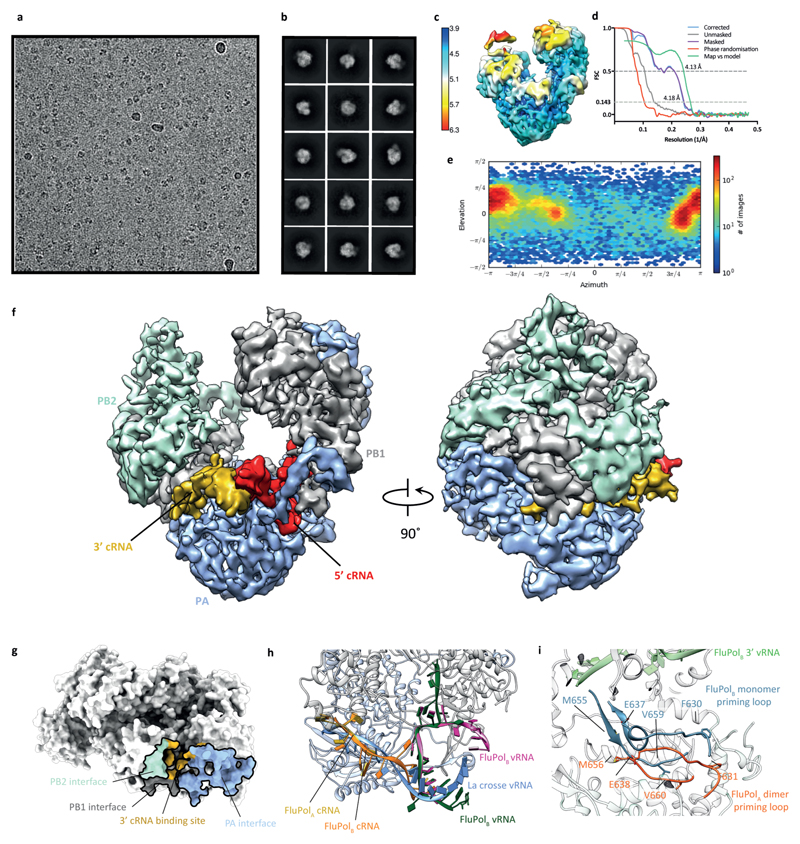
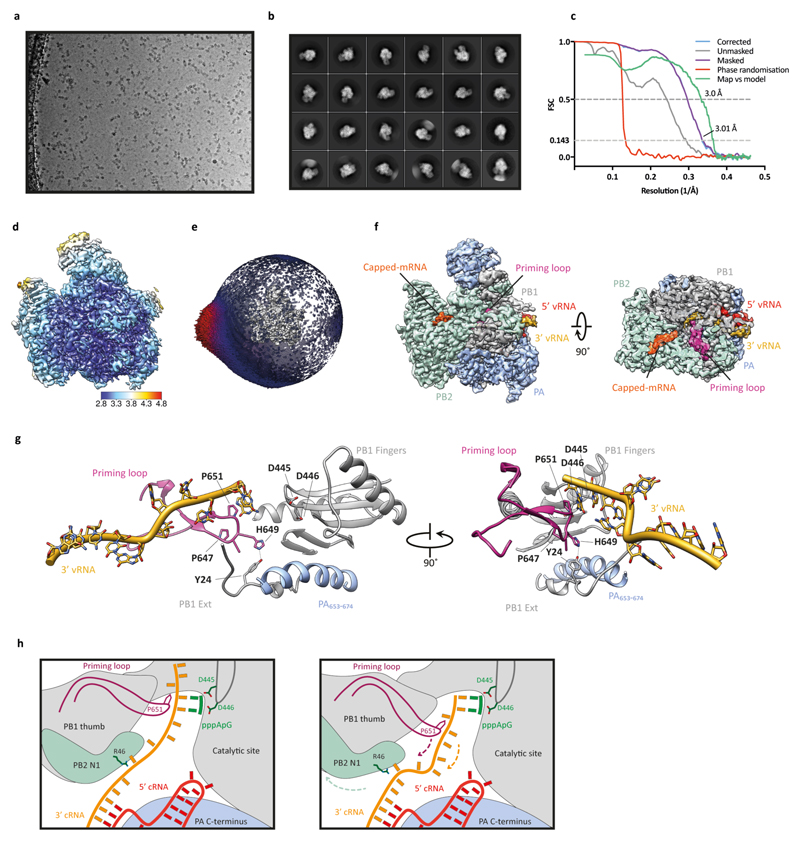
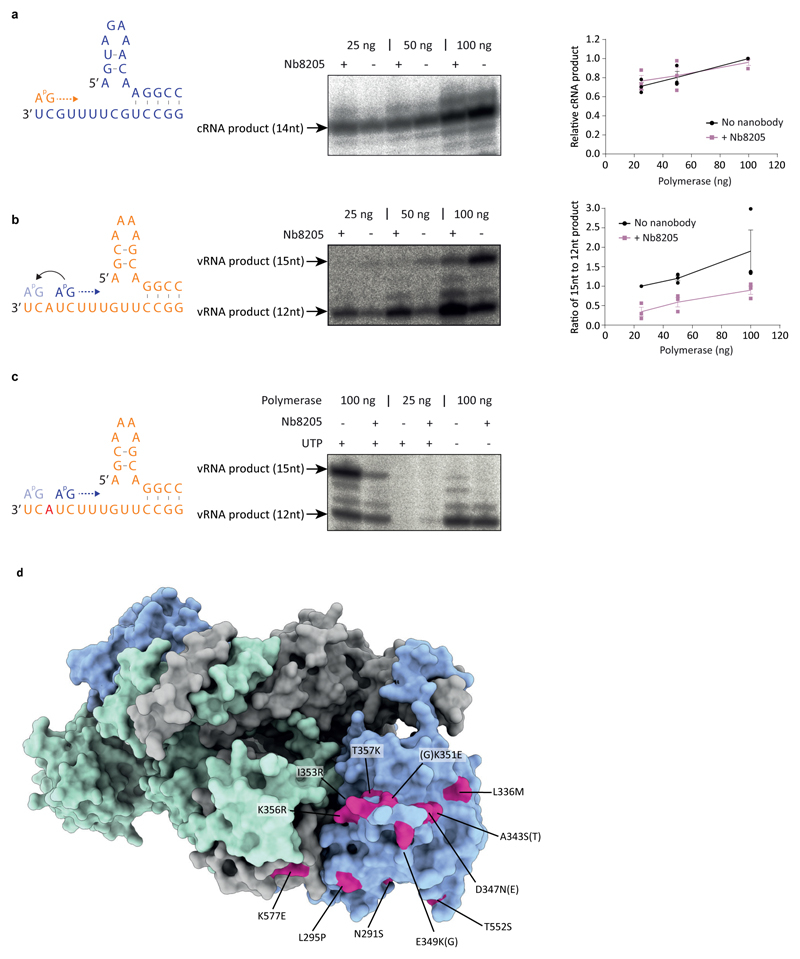
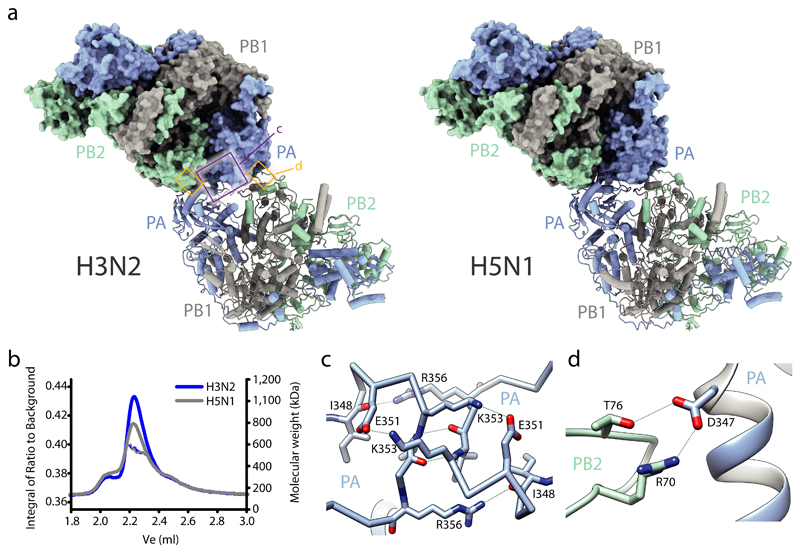
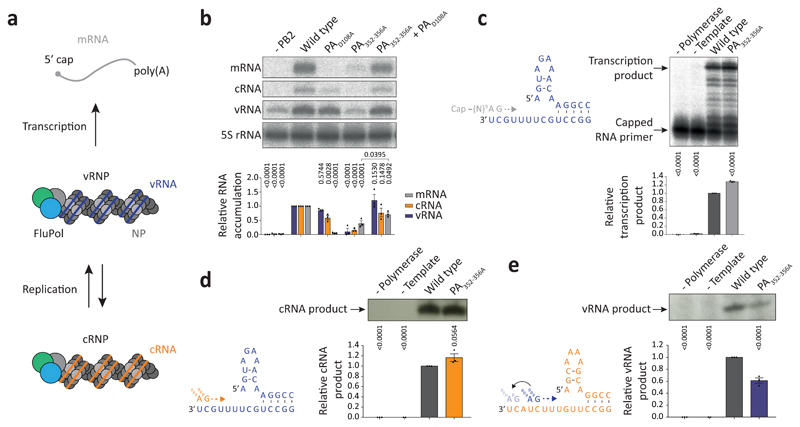
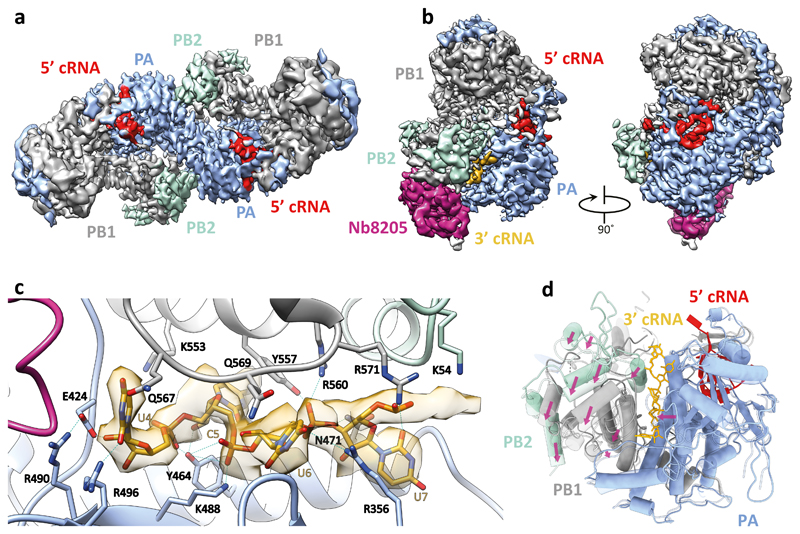
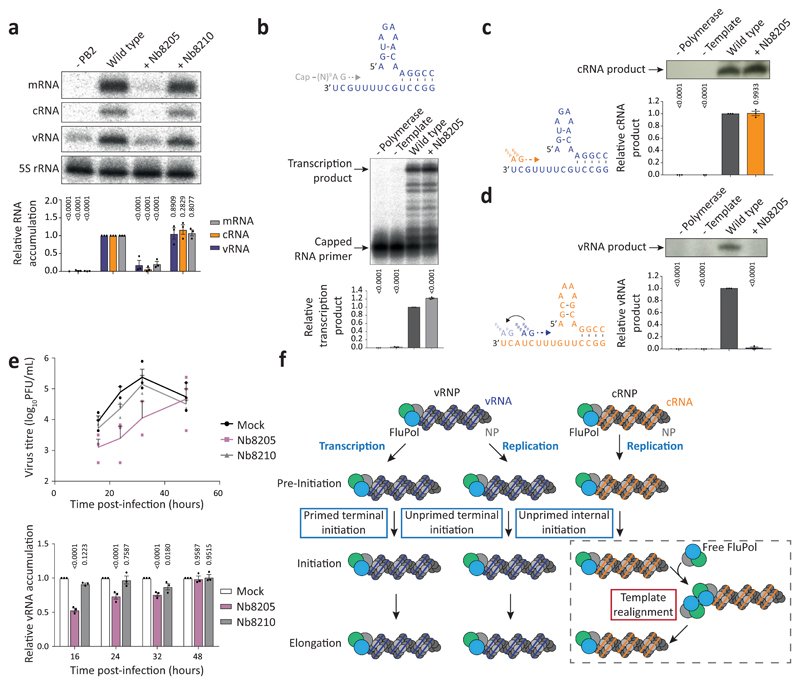
Influenza A viruses are responsible for seasonal epidemics, and pandemics can arise from the transmission of novel zoonotic influenza A viruses to humans1,2. Influenza A viruses contain a segmented negative-sense RNA genome, which is transcribed and replicated by the viral-RNA-dependent RNA polymerase (FluPolA) composed of PB1, PB2 and PA subunits3-5. Although the high-resolution crystal structure of FluPolA of bat influenza A virus has previously been reported6, there are no complete structures available for human and avian FluPolA. Furthermore, the molecular mechanisms of genomic viral RNA (vRNA) replication-which proceeds through a complementary RNA (cRNA) replicative intermediate, and requires oligomerization of the polymerase7-10-remain largely unknown. Here, using crystallography and cryo-electron microscopy, we determine the structures of FluPolA from human influenza A/NT/60/1968 (H3N2) and avian influenza A/duck/Fujian/01/2002 (H5N1) viruses at a resolution of 3.0-4.3 Å, in the presence or absence of a cRNA or vRNA template. In solution, FluPolA forms dimers of heterotrimers through the C-terminal domain of the PA subunit, the thumb subdomain of PB1 and the N1 subdomain of PB2. The cryo-electron microscopy structure of monomeric FluPolA bound to the cRNA template reveals a binding site for the 3' cRNA at the dimer interface. We use a combination of cell-based and in vitro assays to show that the interface of the FluPolA dimer is required for vRNA synthesis during replication of the viral genome. We also show that a nanobody (a single-domain antibody) that interferes with FluPolA dimerization inhibits the synthesis of vRNA and, consequently, inhibits virus replication in infected cells. Our study provides high-resolution structures of medically relevant FluPolA, as well as insights into the replication mechanisms of the viral RNA genome. In addition, our work identifies sites in FluPolA that could be targeted in the development of antiviral drugs.
Conflict of interest statement
Figures


References
-
- Pflug A, Lukarska M, Resa-Infante P, Reich S, Cusack S. Structural insights into RNA synthesis by the influenza virus transcription-replication machine. Virus Res. 2017;234:103–117. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
