

Sargassum fusiforme Fucoidan SP2 Extends the Lifespan of Drosophila melanogaster by Upregulating the Nrf2-Mediated Antioxidant Signaling Pathway
- PMID: 31485301
- PMCID: PMC6710776
- DOI: 10.1155/2019/8918914
Sargassum fusiforme Fucoidan SP2 Extends the Lifespan of Drosophila melanogaster by Upregulating the Nrf2-Mediated Antioxidant Signaling Pathway
Abstract
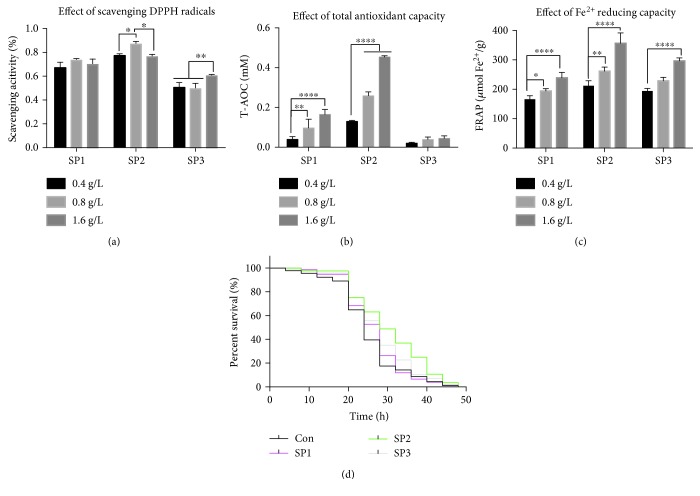
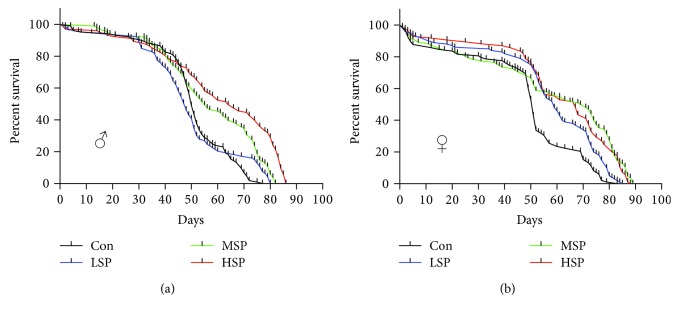
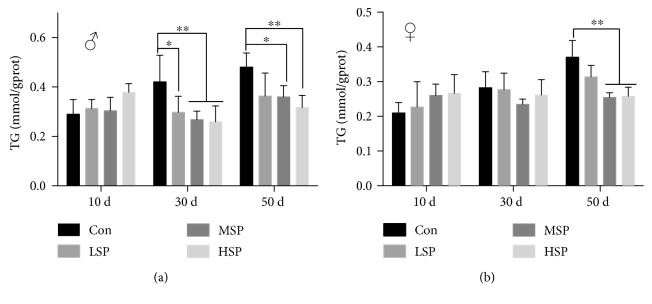
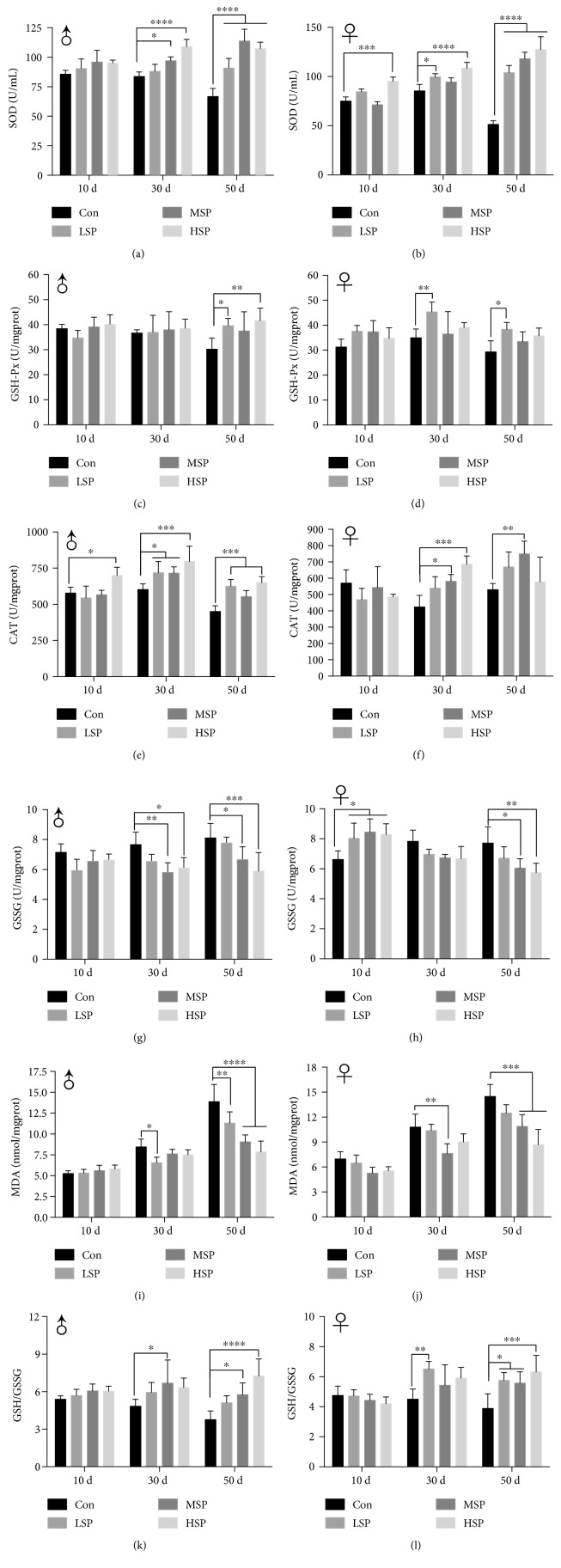
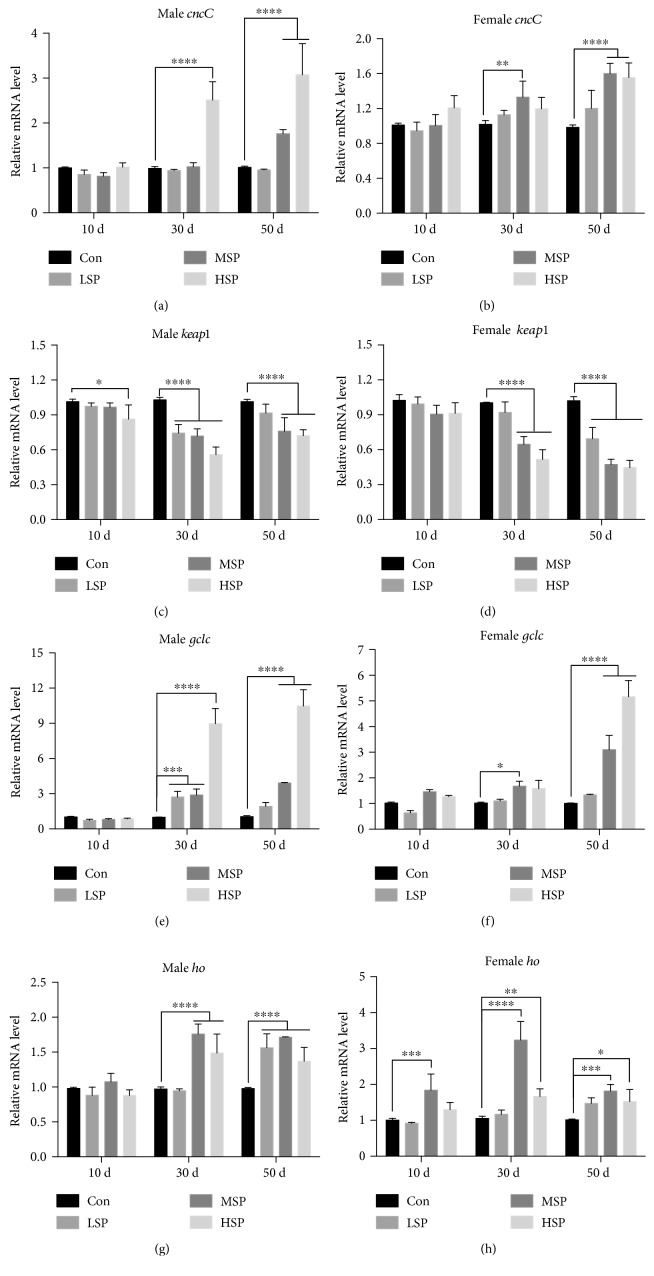
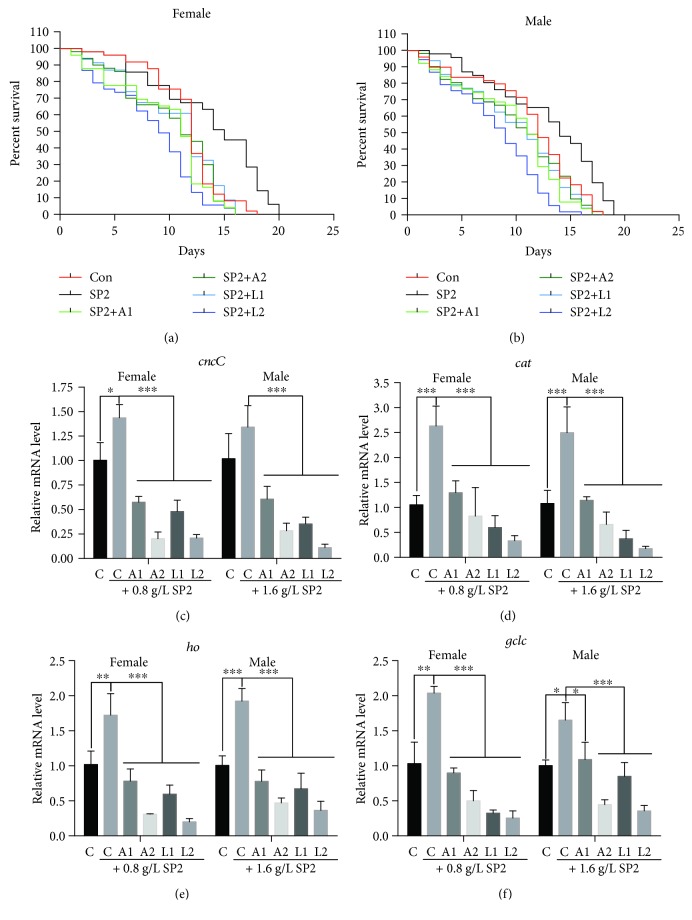
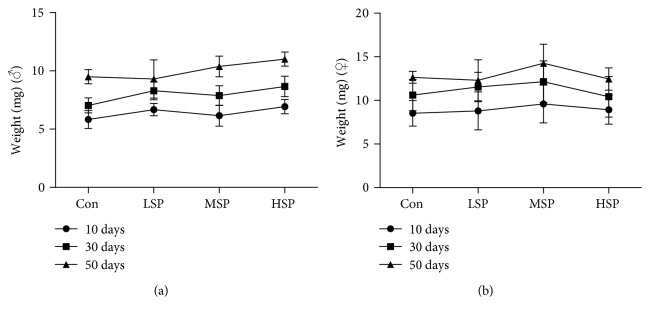
Damage accumulated in the genome and macromolecules is largely attributed to increased oxidative damage and a lack of damage repair in a cell, and this can eventually trigger the process of aging. Alleviating the extent of oxidative damage is therefore considered as a potential way to promote longevity. SFPS, a heteropolysaccharide extracted from the brown alga Sargassum fusiforme, has previously been shown to alleviate oxidative damage during the aging process in mice, but whether SFPS could extend the lifespan of an organism was not demonstrated. Furthermore, the precise component within SFPS that is responsible for the antioxidant activity and the underlying mechanism of such activity was also not resolved. In this study, SP2, a fucoidan derived from SFPS, was shown to exhibit strong antioxidant activity as measured by in vitro radical-scavenging assays. SP2 also improved the survival rate of D. melanogaster subjected to oxidative stress. The flies that were fed with a diet containing SP2 from the time of eclosion displayed significant enhancement in lifespan and reduced accumulation of triglyceride at the old-age stage. In addition, SP2 markedly improved the activities of the antioxidant enzymes, superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GSH-Px) and reduced the contents of the malondialdehyde (MDA) and oxidized glutathione (GSSG) in old flies. Furthermore, SP2 also upregulated the expression levels of the nuclear factor-erythroid-2-like 2 (nfe2l2 or nrf2) and its downstream target genes, accompanied by a dramatic reduction in the expression of kelch-like ECH-associated protein 1 (keap1, a canonical inhibitor of the Nrf2) in old flies. Additional support linking the crucial role of the Nrf2/ARE pathway to the antioxidant effect of SP2 was the relatively high survival rate under heat stress for D. melanogaster individuals receiving SP2 supplement, an effect that was abolished by the inclusion of inhibitors specific for the Nrf2/ARE pathway. Collectively, the results indicated that SP2, a S. fusiforme fucoidan, could promote longevity in D. melanogaster by enhancing the Nrf2-mediated antioxidant signaling pathway during the aging process.
Conflict of interest statement
The authors declare that there are no conflicts of interest regarding the publication of this paper.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
