

Modulation of Tetraspanin 32 (TSPAN32) Expression in T Cell-Mediated Immune Responses and in Multiple Sclerosis
- PMID: 31487788
- PMCID: PMC6770290
- DOI: 10.3390/ijms20184323
Modulation of Tetraspanin 32 (TSPAN32) Expression in T Cell-Mediated Immune Responses and in Multiple Sclerosis
Abstract
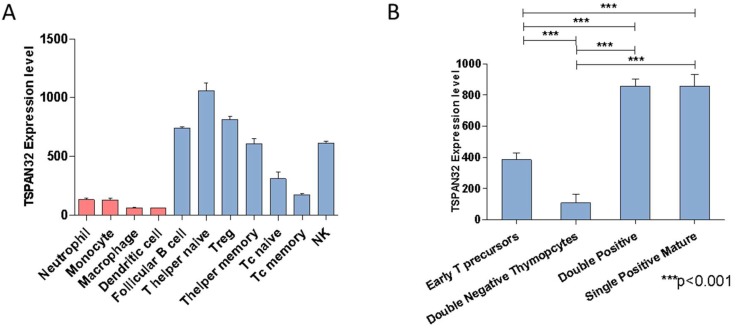
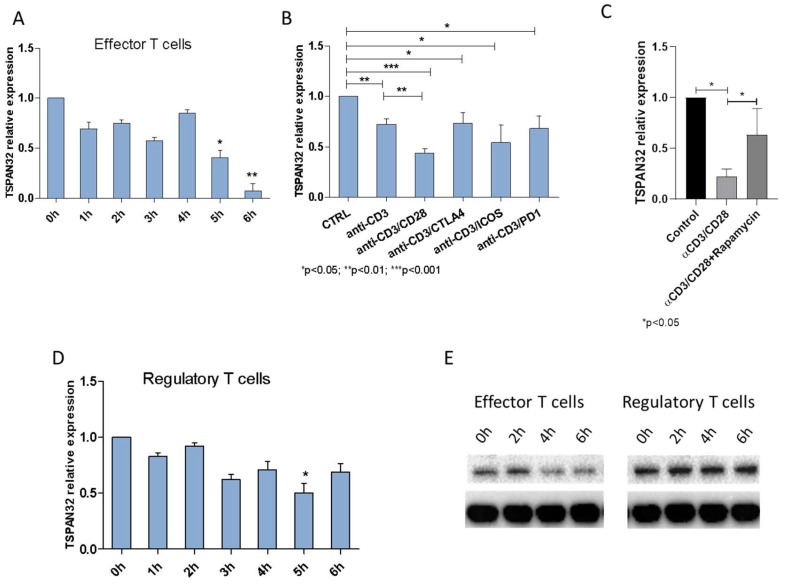
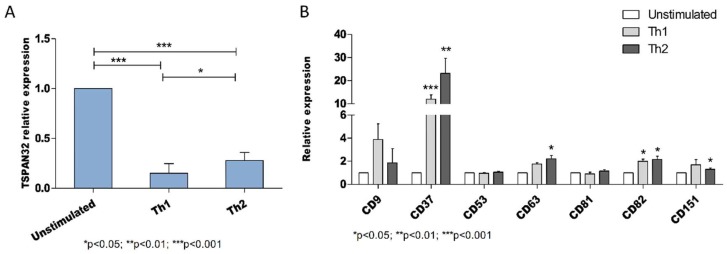
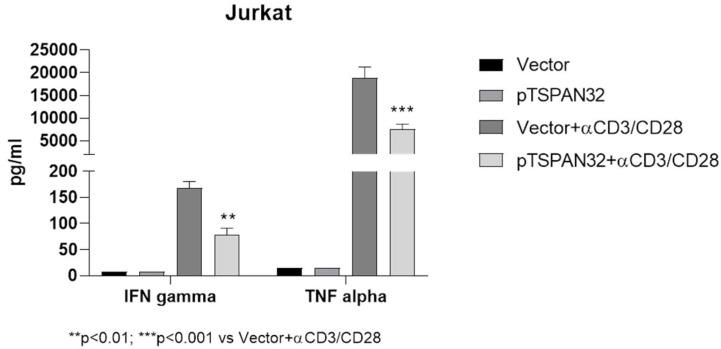
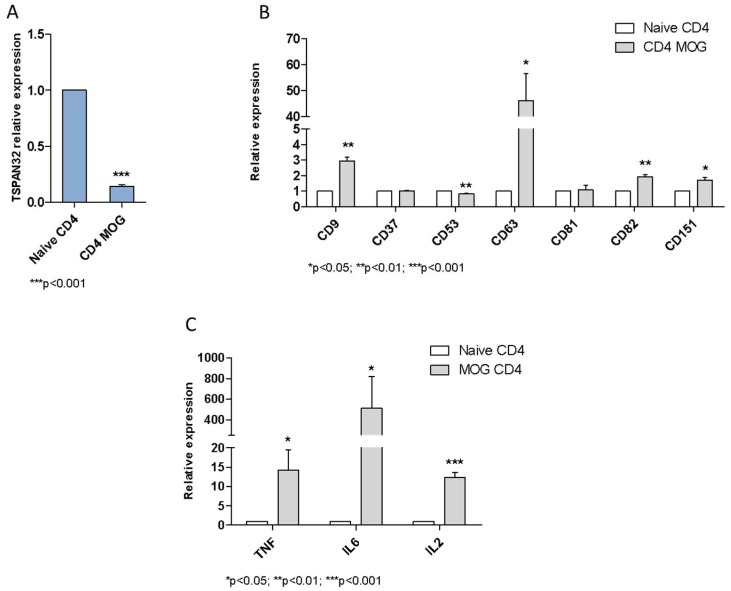
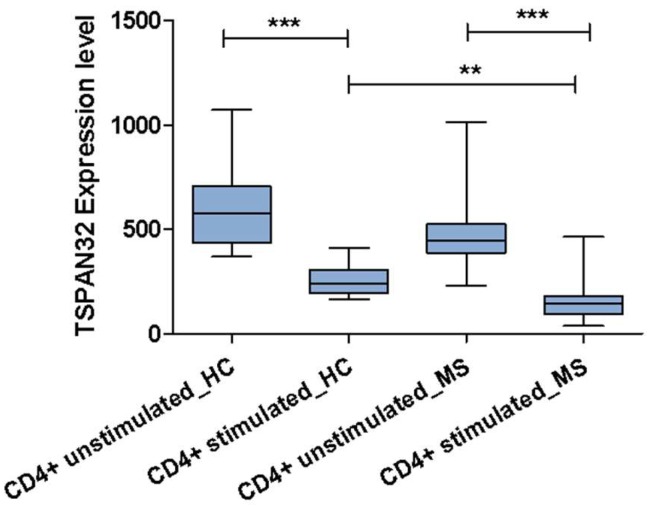
Tetraspanins are a conserved family of proteins involved in a number of biological processes including, cell-cell interactions, fertility, cancer metastasis and immune responses. It has previously been shown that TSPAN32 knockout mice have normal hemopoiesis and B-cell responses, but hyperproliferative T cells. Here, we show that TSPAN32 is expressed at higher levels in the lymphoid lineage as compared to myeloid cells. In vitro activation of T helper cells via anti-CD3/CD28 is associated with a significant downregulation of TSPAN32. Interestingly, engagement of CD3 is sufficient to modulate TSPAN32 expression, and its effect is potentiated by costimulation with anti-CD28, but not anti-CTLA4, -ICOS nor -PD1. Accordingly, we measured the transcriptomic levels of TSPAN32 in polarized T cells under Th1 and Th2 conditions and TSPAN32 resulted significantly reduced as compared with unstimulated cells. On the other hand, in Treg cells, TSPAN32 underwent minor changes upon activation. The in vitro data were finally translated into the context of multiple sclerosis (MS). Encephalitogenic T cells from Myelin Oligodendrocyte Glycoprotein (MOG)-Induced Experimental Autoimmune Encephalomyelitis (EAE) mice showed significantly lower levels of TSPAN32 and increased levels of CD9, CD53, CD82 and CD151. Similarly, in vitro-activated circulating CD4 T cells from MS patients showed lower levels of TSPAN32 as compared with cells from healthy donors. Overall, these data suggest an immunoregulatory role for TSPAN32 in T helper immune response and may represent a target of future immunoregulatory therapies for T cell-mediated autoimmune diseases.
Keywords: T cell responses; TSPAN32; multiple sclerosis; tetraspanins.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Impaired Expression of Tetraspanin 32 (TSPAN32) in Memory T Cells of Patients with Multiple Sclerosis.Brain Sci. 2020 Jan 17;10(1):52. doi: 10.3390/brainsci10010052. Brain Sci. 2020. PMID: 31963428 Free PMC article.
-
Altered Expression of TSPAN32 during B Cell Activation and Systemic Lupus Erythematosus.Genes (Basel). 2021 Jun 18;12(6):931. doi: 10.3390/genes12060931. Genes (Basel). 2021. PMID: 34207245 Free PMC article.
-
Chronological changes of CD4(+) and CD8(+) T cell subsets in the experimental autoimmune encephalomyelitis, a mouse model of multiple sclerosis.Tohoku J Exp Med. 2007 Dec;213(4):329-39. doi: 10.1620/tjem.213.329. Tohoku J Exp Med. 2007. PMID: 18075237
-
T- and B-cell responses to myelin oligodendrocyte glycoprotein in experimental autoimmune encephalomyelitis and multiple sclerosis.Glia. 2001 Nov;36(2):220-34. doi: 10.1002/glia.1111. Glia. 2001. PMID: 11596130 Review.
-
The role of costimulation in autoimmune demyelination.J Neuroimmunol. 2000 Jul 24;107(2):205-15. doi: 10.1016/s0165-5728(00)00230-7. J Neuroimmunol. 2000. PMID: 10854658 Review.
Cited by
-
Effects of Treatment with the Hypomethylating Agent 5-aza-2'-deoxycytidine in Murine Type II Collagen-Induced Arthritis.Pharmaceuticals (Basel). 2019 Nov 27;12(4):174. doi: 10.3390/ph12040174. Pharmaceuticals (Basel). 2019. PMID: 31783688 Free PMC article.
-
TSPAN32 suppresses chronic myeloid leukemia pathogenesis and progression by stabilizing PTEN.Signal Transduct Target Ther. 2023 Mar 1;8(1):90. doi: 10.1038/s41392-022-01290-7. Signal Transduct Target Ther. 2023. PMID: 36854750 Free PMC article.
-
TCF3 and ID3 Regulate TSPAN32 Expression in Burkitt Lymphoma.Scand J Immunol. 2025 Jul;102(1):e70040. doi: 10.1111/sji.70040. Scand J Immunol. 2025. PMID: 40572059 Free PMC article.
-
The State of the Art of Pediatric Multiple Sclerosis.Int J Mol Sci. 2023 May 4;24(9):8251. doi: 10.3390/ijms24098251. Int J Mol Sci. 2023. PMID: 37175954 Free PMC article. Review.
-
Transcriptomic Analysis Reveals Involvement of the Macrophage Migration Inhibitory Factor Gene Network in Duchenne Muscular Dystrophy.Genes (Basel). 2019 Nov 18;10(11):939. doi: 10.3390/genes10110939. Genes (Basel). 2019. PMID: 31752120 Free PMC article.
References
-
- Tarrant J.M., Groom J., Metcalf D., Li R., Borobokas B., Wright M.D., Tarlinton D., Robb L. The absence of Tssc6, a member of the tetraspanin superfamily, does not affect lymphoid development but enhances in vitro T cell proliferative responses. Mol. Cell. Biol. 2002;22:5006–5018. doi: 10.1128/MCB.22.14.5006-5018.2002. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
