

ZCCHC8, the nuclear exosome targeting component, is mutated in familial pulmonary fibrosis and is required for telomerase RNA maturation
- PMID: 31488579
- PMCID: PMC6771387
- DOI: 10.1101/gad.326785.119
ZCCHC8, the nuclear exosome targeting component, is mutated in familial pulmonary fibrosis and is required for telomerase RNA maturation
Abstract
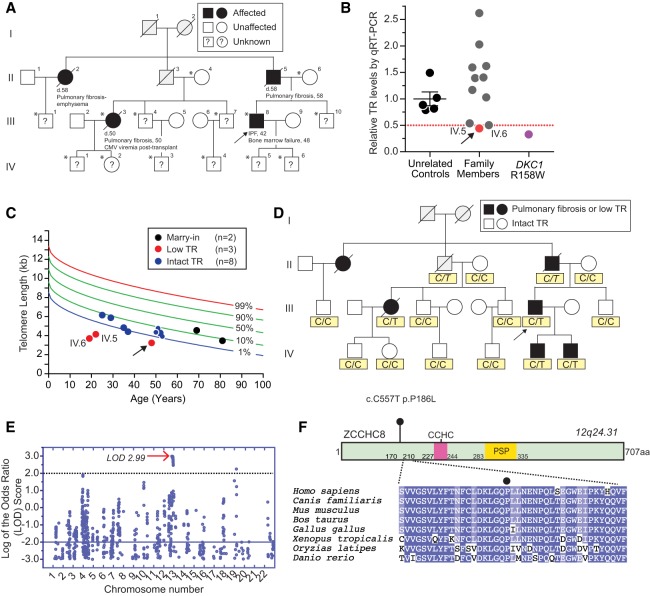
Short telomere syndromes manifest as familial idiopathic pulmonary fibrosis; they are the most common premature aging disorders. We used genome-wide linkage to identify heterozygous loss of function of ZCCHC8, a zinc-knuckle containing protein, as a cause of autosomal dominant pulmonary fibrosis. ZCCHC8 associated with TR and was required for telomerase function. In ZCCHC8 knockout cells and in mutation carriers, genomically extended telomerase RNA (TR) accumulated at the expense of mature TR, consistent with a role for ZCCHC8 in mediating TR 3' end targeting to the nuclear RNA exosome. We generated Zcchc8-null mice and found that heterozygotes, similar to human mutation carriers, had TR insufficiency but an otherwise preserved transcriptome. In contrast, Zcchc8-/- mice developed progressive and fatal neurodevelopmental pathology with features of a ciliopathy. The Zcchc8-/- brain transcriptome was highly dysregulated, showing accumulation and 3' end misprocessing of other low-abundance RNAs, including those encoding cilia components as well as the intronless replication-dependent histones. Our data identify a novel cause of human short telomere syndromes-familial pulmonary fibrosis and uncover nuclear exosome targeting as an essential 3' end maturation mechanism that vertebrate TR shares with replication-dependent histones.
Keywords: RNA processing; ciliopathy; lung disease; nuclear RNA exosome; telomerase RNA.
© 2019 Gable et al.; Published by Cold Spring Harbor Laboratory Press.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous