

Recent Advances on the Structure and Function of RNA Acetyltransferase Kre33/NAT10
- PMID: 31491951
- PMCID: PMC6770127
- DOI: 10.3390/cells8091035
Recent Advances on the Structure and Function of RNA Acetyltransferase Kre33/NAT10
Abstract
Ribosome biogenesis is one of the most energy demanding processes in the cell. In eukaryotes, the main steps of this process occur in the nucleolus and include pre-ribosomal RNA (pre-rRNA) processing, post-transcriptional modifications, and assembly of many non-ribosomal factors and ribosomal proteins in order to form mature and functional ribosomes. In yeast and humans, the nucleolar RNA acetyltransferase Kre33/NAT10 participates in different maturation events, such as acetylation and processing of 18S rRNA, and assembly of the 40S ribosomal subunit. Here, we review the structural and functional features of Kre33/NAT10 RNA acetyltransferase, and we underscore the importance of this enzyme in ribosome biogenesis, as well as in acetylation of non-ribosomal targets. We also report on the role of human NAT10 in Hutchinson-Gilford progeria syndrome.
Keywords: Hutchinson–Gilford progeria syndrome; Kre33; NAT10; SSU processome; nuclear localization signal; nuclear pore complex; nucleolar localization signal; post-transcriptional modifications; ribosome biogenesis.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures
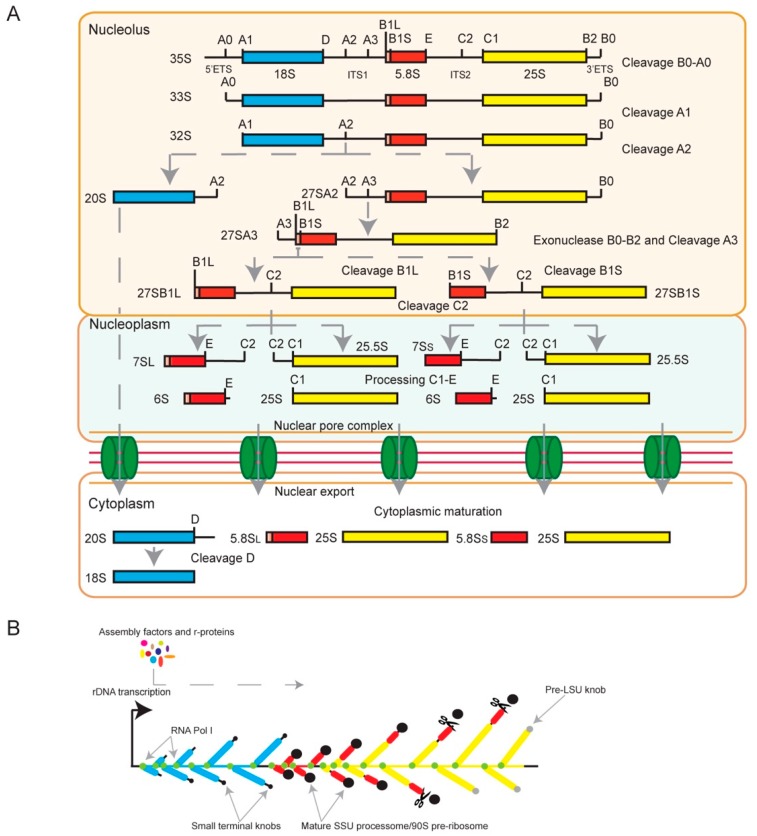
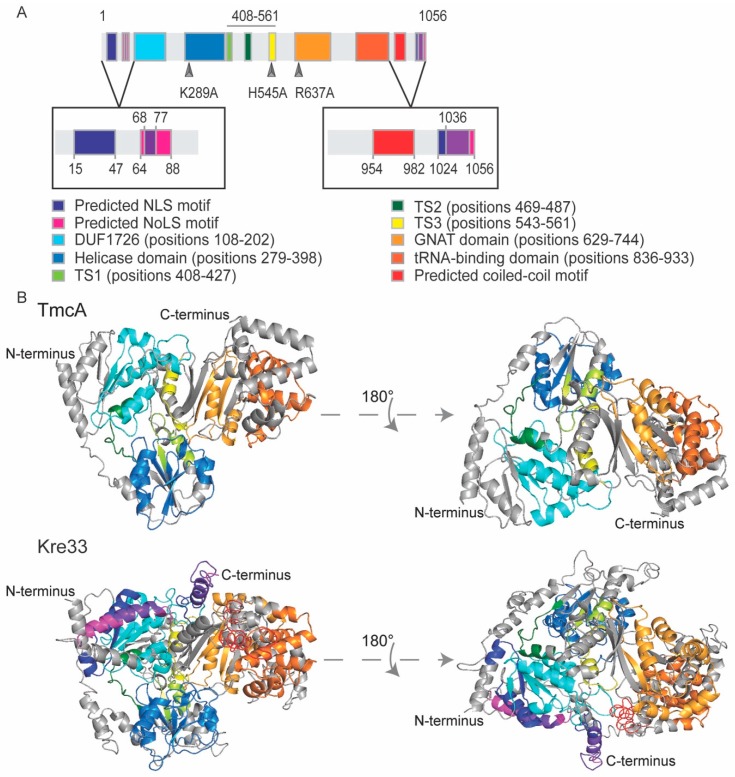
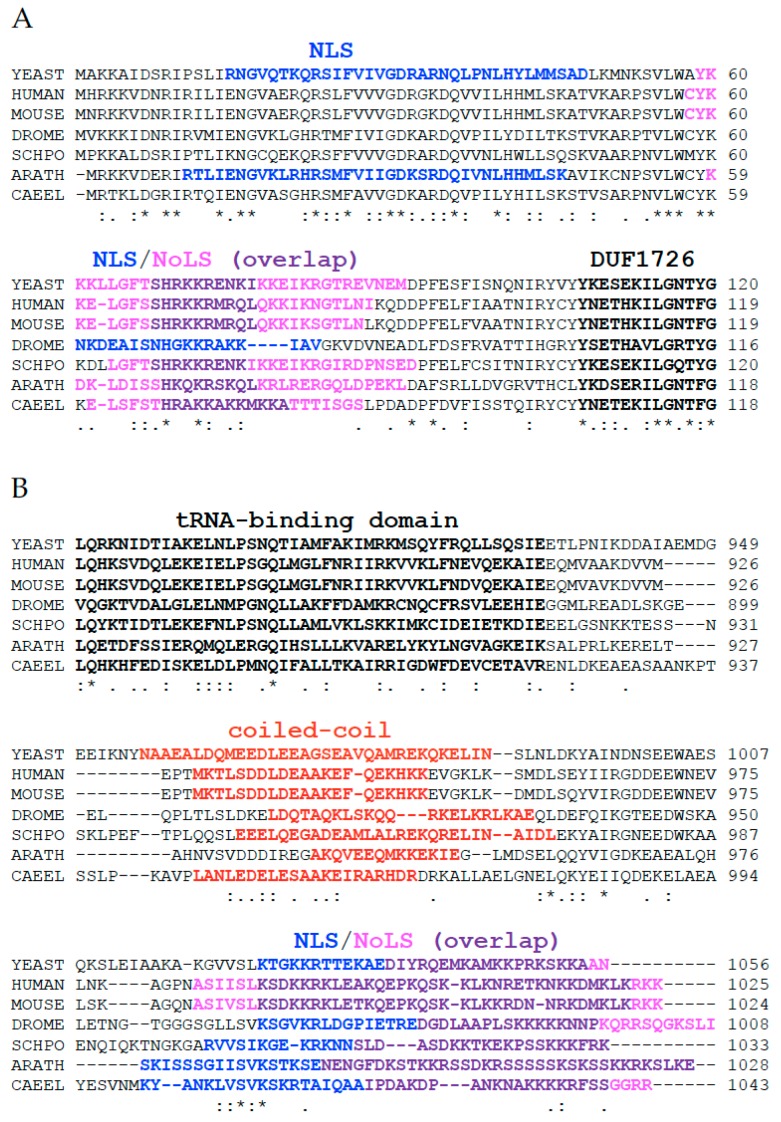
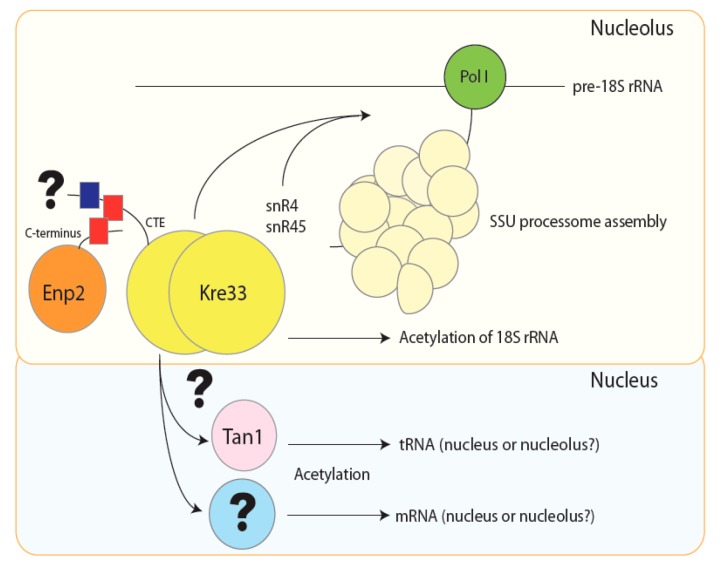
Similar articles
-
Yeast Kre33 and human NAT10 are conserved 18S rRNA cytosine acetyltransferases that modify tRNAs assisted by the adaptor Tan1/THUMPD1.Nucleic Acids Res. 2015 Feb 27;43(4):2242-58. doi: 10.1093/nar/gkv075. Epub 2015 Feb 4. Nucleic Acids Res. 2015. PMID: 25653167 Free PMC article.
-
A single acetylation of 18 S rRNA is essential for biogenesis of the small ribosomal subunit in Saccharomyces cerevisiae.J Biol Chem. 2014 Sep 19;289(38):26201-26212. doi: 10.1074/jbc.M114.593996. Epub 2014 Aug 1. J Biol Chem. 2014. PMID: 25086048 Free PMC article.
-
Advances of NAT10 in diseases: insights from dual properties as protein and RNA acetyltransferase.Cell Biol Toxicol. 2024 Dec 27;41(1):17. doi: 10.1007/s10565-024-09962-6. Cell Biol Toxicol. 2024. PMID: 39725720 Free PMC article. Review.
-
Human NAT10 is an ATP-dependent RNA acetyltransferase responsible for N4-acetylcytidine formation in 18 S ribosomal RNA (rRNA).J Biol Chem. 2014 Dec 26;289(52):35724-30. doi: 10.1074/jbc.C114.602698. Epub 2014 Nov 19. J Biol Chem. 2014. PMID: 25411247 Free PMC article.
-
A Review of Research on the Mechanism of Tumor Regulation by N-Acetyltransferase 10.Discov Med. 2024 Jul;36(186):1334-1344. doi: 10.24976/Discov.Med.202436186.123. Discov Med. 2024. PMID: 39054704 Review.
Cited by
-
NAT10 Promotes Osteogenic Differentiation of Mesenchymal Stem Cells by Mediating N4-Acetylcytidine Modification of Gremlin 1.Stem Cells Int. 2021 Apr 12;2021:8833527. doi: 10.1155/2021/8833527. eCollection 2021. Stem Cells Int. 2021. PMID: 33953754 Free PMC article.
-
N-acetyltransferase 10 promotes cutaneous wound repair via the NF-κB-IL-6 axis.Cell Death Discov. 2023 Aug 29;9(1):324. doi: 10.1038/s41420-023-01628-2. Cell Death Discov. 2023. PMID: 37644005 Free PMC article.
-
Ac4C Enhances the Translation Efficiency of Vegfa mRNA and Mediates Central Sensitization in Spinal Dorsal Horn in Neuropathic Pain.Adv Sci (Weinh). 2023 Dec;10(35):e2303113. doi: 10.1002/advs.202303113. Epub 2023 Oct 25. Adv Sci (Weinh). 2023. PMID: 37877615 Free PMC article.
-
Poly(ADP-ribosyl)ation of acetyltransferase NAT10 by PARP1 is required for its nucleoplasmic translocation and function in response to DNA damage.Cell Commun Signal. 2022 Aug 19;20(1):127. doi: 10.1186/s12964-022-00932-1. Cell Commun Signal. 2022. PMID: 35986334 Free PMC article.
-
ac4C acetylation regulates mRNA stability and translation efficiency in osteosarcoma.Heliyon. 2023 Jun 8;9(6):e17103. doi: 10.1016/j.heliyon.2023.e17103. eCollection 2023 Jun. Heliyon. 2023. PMID: 37484432 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
