

Universality of fold-encoded localized vibrations in enzymes
- PMID: 31492876
- PMCID: PMC6731342
- DOI: 10.1038/s41598-019-48905-8
Universality of fold-encoded localized vibrations in enzymes
Abstract
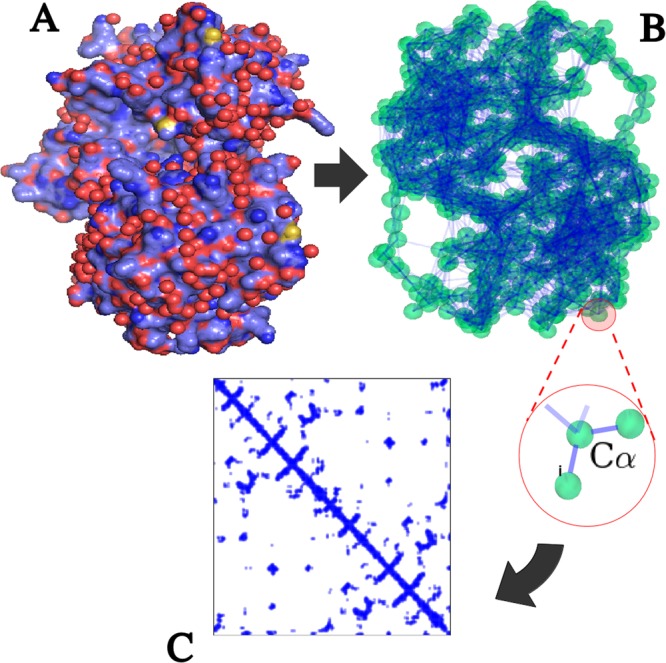
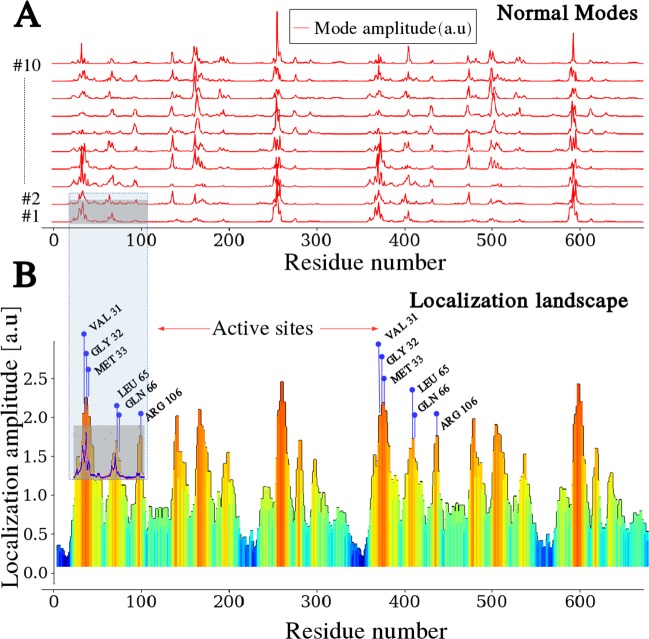
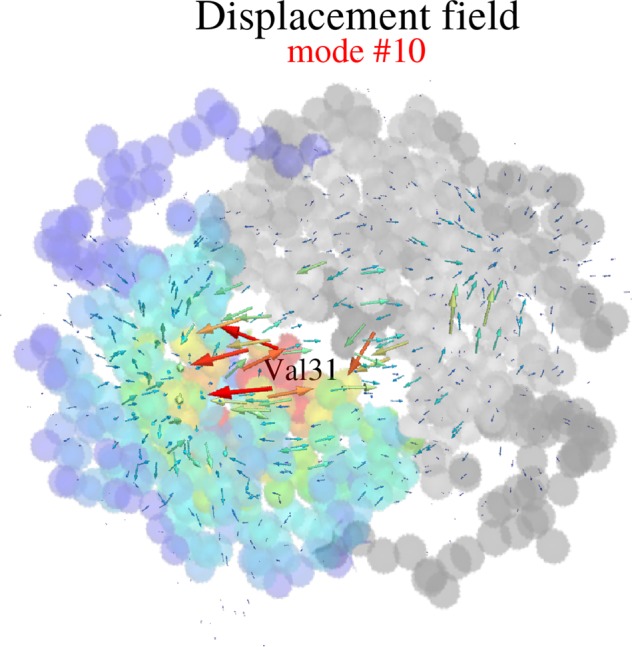
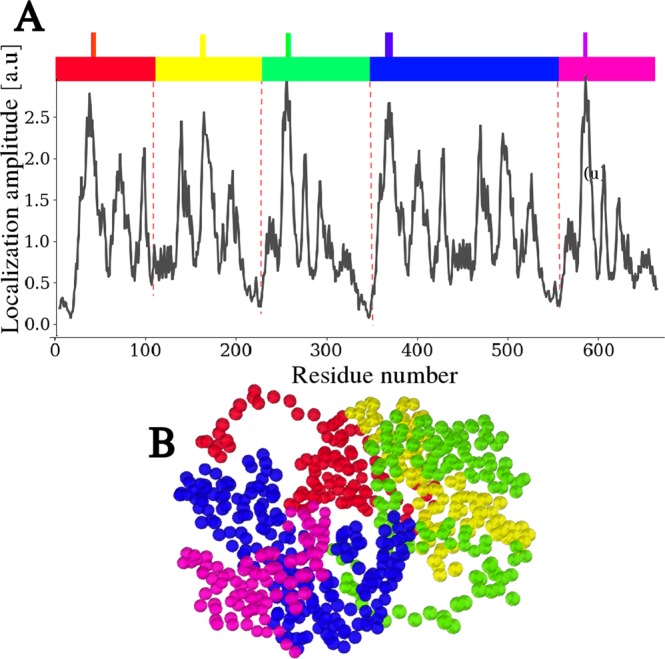
Enzymes speed up biochemical reactions at the core of life by as much as 15 orders of magnitude. Yet, despite considerable advances, the fine dynamical determinants at the microscopic level of their catalytic proficiency are still elusive. In this work, we use a powerful mathematical approach to show that rate-promoting vibrations in the picosecond range, specifically encoded in the 3D protein structure, are localized vibrations optimally coupled to the chemical reaction coordinates at the active site. Remarkably, our theory also exposes an hithertho unknown deep connection between the unique localization fingerprint and a distinct partition of the 3D fold into independent, foldspanning subdomains that govern long-range communication. The universality of these features is demonstrated on a pool of more than 900 enzyme structures, comprising a total of more than 10,000 experimentally annotated catalytic sites. Our theory provides a unified microscopic rationale for the subtle structure-dynamics-function link in proteins.
Conflict of interest statement
The authors declare no competing interests.
Figures




Similar articles
-
Protein dynamics and enzymatic chemical barrier passage.J Phys Chem B. 2011 Dec 29;115(51):15147-58. doi: 10.1021/jp207876k. Epub 2011 Nov 15. J Phys Chem B. 2011. PMID: 22031954 Free PMC article.
-
The dynamical nature of enzymatic catalysis.Acc Chem Res. 2015 Feb 17;48(2):407-13. doi: 10.1021/ar5002928. Epub 2014 Dec 24. Acc Chem Res. 2015. PMID: 25539144 Free PMC article.
-
Targeting a Rate-Promoting Vibration with an Allosteric Mediator in Lactate Dehydrogenase.J Phys Chem Lett. 2016 Jul 7;7(13):2591-6. doi: 10.1021/acs.jpclett.6b01209. Epub 2016 Jun 24. J Phys Chem Lett. 2016. PMID: 27327209 Free PMC article.
-
Low-Frequency Protein Motions Coupled to Catalytic Sites.Annu Rev Phys Chem. 2020 Apr 20;71:267-288. doi: 10.1146/annurev-physchem-050317-014308. Annu Rev Phys Chem. 2020. PMID: 32312192 Review.
-
Hybrid schemes based on quantum mechanics/molecular mechanics simulations goals to success, problems, and perspectives.Adv Protein Chem Struct Biol. 2011;85:81-142. doi: 10.1016/B978-0-12-386485-7.00003-X. Adv Protein Chem Struct Biol. 2011. PMID: 21920322 Review.
Cited by
-
Phonon-assisted electron-proton transfer in [FeFe] hydrogenases: Topological role of clusters.Biophys J. 2023 Apr 18;122(8):1557-1567. doi: 10.1016/j.bpj.2023.03.027. Epub 2023 Mar 23. Biophys J. 2023. PMID: 36960530 Free PMC article.
-
Utilizing Molecular Dynamics Simulations, Machine Learning, Cryo-EM, and NMR Spectroscopy to Predict and Validate Protein Dynamics.Int J Mol Sci. 2024 Sep 8;25(17):9725. doi: 10.3390/ijms25179725. Int J Mol Sci. 2024. PMID: 39273672 Free PMC article. Review.
-
The physical origin of rate promoting vibrations in enzymes revealed by structural rigidity.Sci Rep. 2020 Oct 15;10(1):17465. doi: 10.1038/s41598-020-74439-5. Sci Rep. 2020. PMID: 33060716 Free PMC article.
-
Energy Bilocalization Effect and the Emergence of Molecular Functions in Proteins.Front Mol Biosci. 2021 Dec 23;8:736376. doi: 10.3389/fmolb.2021.736376. eCollection 2021. Front Mol Biosci. 2021. PMID: 35004841 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
