

Juxta-membrane S-acylation of plant receptor-like kinases is likely fortuitous and does not necessarily impact upon function
- PMID: 31492958
- PMCID: PMC6731221
- DOI: 10.1038/s41598-019-49302-x
Juxta-membrane S-acylation of plant receptor-like kinases is likely fortuitous and does not necessarily impact upon function
Abstract
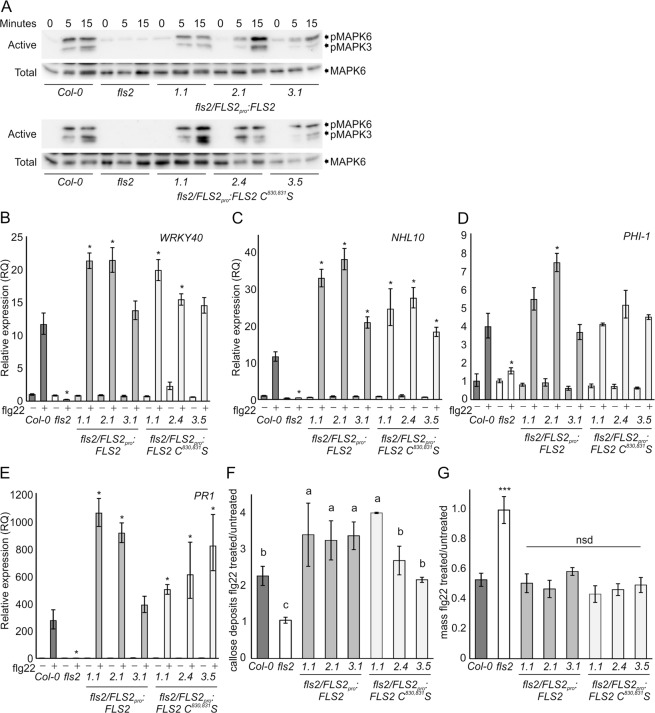
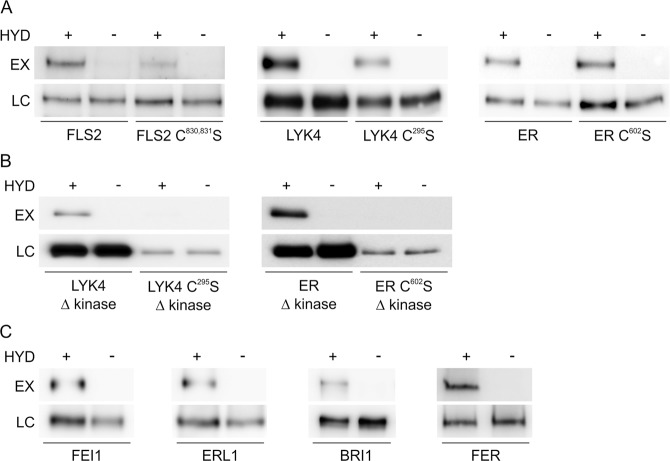
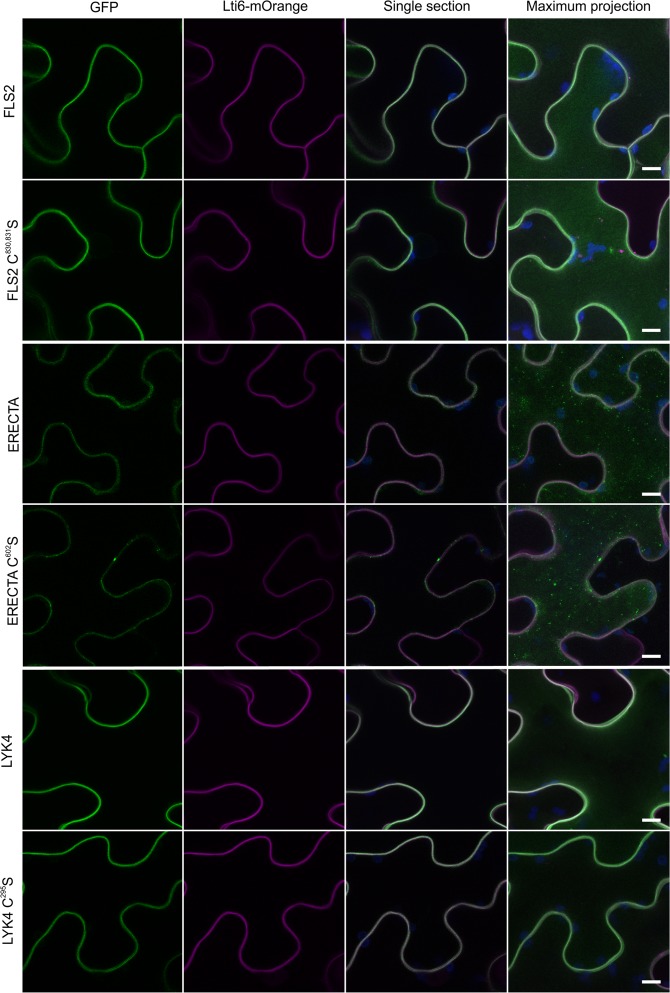
S-acylation is a common post-translational modification of membrane protein cysteine residues with many regulatory roles. S-acylation adjacent to transmembrane domains has been described in the literature as affecting diverse protein properties including turnover, trafficking and microdomain partitioning. However, all of these data are derived from mammalian and yeast systems. Here we examine the role of S-acylation adjacent to the transmembrane domain of the plant pathogen perceiving receptor-like kinase FLS2. Surprisingly, S-acylation of FLS2 adjacent to the transmembrane domain is not required for either FLS2 trafficking or signalling function. Expanding this analysis to the wider plant receptor-like kinase family we find that S-acylation adjacent to receptor-like kinase domains is common, affecting ~25% of Arabidopsis receptor-like kinases, but poorly conserved between orthologues through evolution. This suggests that S-acylation of receptor-like kinases at this site is likely the result of chance mutation leading to cysteine occurrence. As transmembrane domains followed by cysteine residues are common motifs for S-acylation to occur, and many S-acyl transferases appear to have lax substrate specificity, we propose that many receptor-like kinases are fortuitously S-acylated once chance mutation has introduced a cysteine at this site. Interestingly some receptor-like kinases show conservation of S-acylation sites between orthologues suggesting that S-acylation has come to play a role and has been positively selected for during evolution. The most notable example of this is in the ERECTA-like family where S-acylation of ERECTA adjacent to the transmembrane domain occurs in all ERECTA orthologues but not in the parental ERECTA-like clade. This suggests that ERECTA S-acylation occurred when ERECTA emerged during the evolution of angiosperms and may have contributed to the neo-functionalisation of ERECTA from ERECTA-like proteins.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
S-acylation stabilizes ligand-induced receptor kinase complex formation during plant pattern-triggered immune signaling.Curr Biol. 2023 Apr 24;33(8):1588-1596.e6. doi: 10.1016/j.cub.2023.02.065. Epub 2023 Mar 15. Curr Biol. 2023. PMID: 36924767
-
A proteomic approach identifies many novel palmitoylated proteins in Arabidopsis.New Phytol. 2013 Feb;197(3):805-814. doi: 10.1111/nph.12077. Epub 2012 Dec 17. New Phytol. 2013. PMID: 23252521
-
A Cysteine-Rich Protein Kinase Associates with a Membrane Immune Complex and the Cysteine Residues Are Required for Cell Death.Plant Physiol. 2017 Jan;173(1):771-787. doi: 10.1104/pp.16.01404. Epub 2016 Nov 16. Plant Physiol. 2017. PMID: 27852951 Free PMC article.
-
Mapping FLS2 function to structure: LRRs, kinase and its working bits.Protoplasma. 2013 Jun;250(3):671-81. doi: 10.1007/s00709-012-0459-6. Epub 2012 Oct 5. Protoplasma. 2013. PMID: 23053766 Review.
-
Diverse roles of ERECTA family genes in plant development.J Integr Plant Biol. 2013 Dec;55(12):1238-50. doi: 10.1111/jipb.12108. Epub 2013 Oct 30. J Integr Plant Biol. 2013. PMID: 24016315 Review.
Cited by
-
Arabidopsis class A S-acyl transferases modify the pollen receptors LIP1 and PRK1 to regulate pollen tube guidance.Plant Cell. 2024 Sep 3;36(9):3419-3434. doi: 10.1093/plcell/koae109. Plant Cell. 2024. PMID: 38635962 Free PMC article.
-
An atlas of Arabidopsis protein S-acylation reveals its widespread role in plant cell organization and function.Nat Plants. 2022 Jun;8(6):670-681. doi: 10.1038/s41477-022-01164-4. Epub 2022 Jun 9. Nat Plants. 2022. PMID: 35681017 Free PMC article.
-
Uptake of oomycete RXLR effectors into host cells by clathrin-mediated endocytosis.Plant Cell. 2023 Jun 26;35(7):2504-2526. doi: 10.1093/plcell/koad069. Plant Cell. 2023. PMID: 36911990 Free PMC article.
-
Ligand recognition and signal transduction by lectin receptor-like kinases in plant immunity.Front Plant Sci. 2023 Jun 15;14:1201805. doi: 10.3389/fpls.2023.1201805. eCollection 2023. Front Plant Sci. 2023. PMID: 37396638 Free PMC article. Review.
-
Free Radicals Mediated Redox Signaling in Plant Stress Tolerance.Life (Basel). 2023 Jan 10;13(1):204. doi: 10.3390/life13010204. Life (Basel). 2023. PMID: 36676153 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
